Das könnte Ihnen auch gefallen
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (589)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5795)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1091)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Truth About Climate ChangeDokument12 SeitenThe Truth About Climate ChangeBob Berwyn100% (9)
- Colace (Docusate Sodium)Dokument1 SeiteColace (Docusate Sodium)E100% (1)
- 20170915-Concepts Uk Future c2 JCN 2 17 PDFDokument64 Seiten20170915-Concepts Uk Future c2 JCN 2 17 PDFKebede MichaelNoch keine Bewertungen
- Bavarian FiresDokument11 SeitenBavarian FiresBob BerwynNoch keine Bewertungen
- M. Ionita (Monica - Ionita@awi - De), K. Grosfeld, G. Lohmann and P. ScholzDokument1 SeiteM. Ionita (Monica - Ionita@awi - De), K. Grosfeld, G. Lohmann and P. ScholzBob BerwynNoch keine Bewertungen
- Entry Into Force of Pa-1Dokument8 SeitenEntry Into Force of Pa-1Bob BerwynNoch keine Bewertungen
- For Us, So-Called Subsistence Activity Is Far More Than Subsistence. Hunting Is More Than Food On The Table. It Is A Fundamental Part of Who We Are.Dokument5 SeitenFor Us, So-Called Subsistence Activity Is Far More Than Subsistence. Hunting Is More Than Food On The Table. It Is A Fundamental Part of Who We Are.Bob BerwynNoch keine Bewertungen
- Eakin Et Al 2016 Reef-Encounter-2014-17 BleachingDokument7 SeitenEakin Et Al 2016 Reef-Encounter-2014-17 BleachingBob BerwynNoch keine Bewertungen
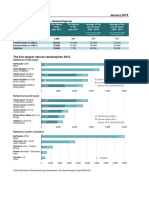
- Munich Re Overview Natural Catastrophes 2015 1Dokument1 SeiteMunich Re Overview Natural Catastrophes 2015 1Bob BerwynNoch keine Bewertungen
- Eakin Et Al 2016 Reef-Encounter-2014-17 BleachingDokument7 SeitenEakin Et Al 2016 Reef-Encounter-2014-17 BleachingBob BerwynNoch keine Bewertungen
- Fracking Setback AnalysisDokument20 SeitenFracking Setback AnalysisBob BerwynNoch keine Bewertungen
- Nepa Final GHG Guidance-1Dokument34 SeitenNepa Final GHG Guidance-1Bob BerwynNoch keine Bewertungen
- OIG Response Letter Chairman Smith Summit 04-29-2016 (3) (2) (1) (1) (1) (1) - 2Dokument3 SeitenOIG Response Letter Chairman Smith Summit 04-29-2016 (3) (2) (1) (1) (1) (1) - 2Bob BerwynNoch keine Bewertungen
- SH9 Public Hearing Display Boards 072914Dokument21 SeitenSH9 Public Hearing Display Boards 072914Bob BerwynNoch keine Bewertungen
- Federal Court Wolverine Endangered Species DecisonDokument85 SeitenFederal Court Wolverine Endangered Species DecisonBob BerwynNoch keine Bewertungen
- CO Fracking LawsuitDokument7 SeitenCO Fracking LawsuitBob BerwynNoch keine Bewertungen
- Methane ForumDokument2 SeitenMethane ForumBob BerwynNoch keine Bewertungen
- Arkansas DispensariesDokument6 SeitenArkansas DispensariesAdam ByrdNoch keine Bewertungen
- AFST - Oct 17Dokument9 SeitenAFST - Oct 17kimkimNoch keine Bewertungen
- MOI Lesson PlanDokument4 SeitenMOI Lesson PlanPreeti KumariNoch keine Bewertungen
- Zachary Evans ResumeDokument2 SeitenZachary Evans Resumeapi-248576943Noch keine Bewertungen
- CFA Level I - Timetable (August 2021 Exam) (V2)Dokument1 SeiteCFA Level I - Timetable (August 2021 Exam) (V2)Via Commerce Sdn BhdNoch keine Bewertungen
- n1 BrochureDokument4 Seitenn1 BrochureMatt JaskulskiNoch keine Bewertungen
- Food Stuff From List CompaniesDokument3 SeitenFood Stuff From List CompaniesListonNoch keine Bewertungen
- 12th Commerce Study Material English MediumDokument97 Seiten12th Commerce Study Material English MediumDeepak ajar100% (1)
- A Three-Stage Layer-Based Heuristic To Solve The 3D Bin-Packing ProblemDokument7 SeitenA Three-Stage Layer-Based Heuristic To Solve The 3D Bin-Packing ProblemEder NingunoNoch keine Bewertungen
- Problem Solving 2Dokument20 SeitenProblem Solving 2Yeppeudda100% (1)
- 01 AR PFMEA - TemplateDokument3 Seiten01 AR PFMEA - TemplateAndrew DoNoch keine Bewertungen
- 4 Insurance Sales Incentive Program Ideas That WorkDokument3 Seiten4 Insurance Sales Incentive Program Ideas That Workgeorge akamaNoch keine Bewertungen
- NewJaisa Corporate ProfileDokument14 SeitenNewJaisa Corporate ProfileCRAZY ಕನ್ನಡಿಗNoch keine Bewertungen
- Bengaluru BDA RMP 2031 PD Index MapDokument206 SeitenBengaluru BDA RMP 2031 PD Index MapmyskmeNoch keine Bewertungen
- Sustainable Transportation and Electric VehiclesDokument17 SeitenSustainable Transportation and Electric Vehicless131744Noch keine Bewertungen
- Knowledge RepresentationDokument14 SeitenKnowledge RepresentationGonibala LandyNoch keine Bewertungen
- Vietnam Maritime UniversityDokument36 SeitenVietnam Maritime UniversitySơn BáchNoch keine Bewertungen
- LM5 RIOL Lite DatasheetDokument12 SeitenLM5 RIOL Lite DatasheetStilux VoltNoch keine Bewertungen
- Sun OBPDokument30 SeitenSun OBPgoyaltcsNoch keine Bewertungen
- Examen EESDokument16 SeitenExamen EESManu LlacsaNoch keine Bewertungen
- Marketing Plan Rental LaptopDokument15 SeitenMarketing Plan Rental LaptopDany AkbarNoch keine Bewertungen
- Oc Ik 100008Dokument1 SeiteOc Ik 100008Jagjeet SinghNoch keine Bewertungen
- 29102015000000B - Boehler DCMS-IG - SWDokument1 Seite29102015000000B - Boehler DCMS-IG - SWErdinc BayatNoch keine Bewertungen
- UntitledDokument42 SeitenUntitledDiana JacquelineNoch keine Bewertungen
- No TitleDokument6 SeitenNo Titlehwh8w6jmzsNoch keine Bewertungen
- Production Technology of Cow PeaDokument20 SeitenProduction Technology of Cow PeaMUHAMMAD AQEELNoch keine Bewertungen
- SosDokument6 SeitenSosVikramNoch keine Bewertungen
- Fpso Revamp For A Marginal Field Ea-SpeDokument34 SeitenFpso Revamp For A Marginal Field Ea-Spe고병석Noch keine Bewertungen