Das könnte Ihnen auch gefallen
- Structura Acizi Nucleici - Curs GeneralDokument127 SeitenStructura Acizi Nucleici - Curs GeneralJoy CarsonNoch keine Bewertungen
- Absorbtia IntestinalaDokument16 SeitenAbsorbtia IntestinalaMădălina Elena AndrişoaeiNoch keine Bewertungen
- Genetica AMG IDokument6 SeitenGenetica AMG IMaria Claudia SelariuNoch keine Bewertungen
- Curs 2 Structura Acizilor Nucleici, ProprietatiDokument48 SeitenCurs 2 Structura Acizilor Nucleici, ProprietatiionutNoch keine Bewertungen
- Teme MicrobiologieDokument4 SeitenTeme MicrobiologieDiana DiaconescuNoch keine Bewertungen
- Curs 11 Pti+leucemii Si HemofiliiDokument44 SeitenCurs 11 Pti+leucemii Si HemofiliiVioleta TudoseNoch keine Bewertungen
- Genetica CursDokument4 SeitenGenetica CursRonaldo LazarNoch keine Bewertungen
- Noţiuni de CitologieDokument8 SeitenNoţiuni de CitologieAnaMihaelaManguNoch keine Bewertungen
- BiochimieDokument5 SeitenBiochimiebluedolphin7Noch keine Bewertungen
- Psihologia MedicamentuluiDokument2 SeitenPsihologia MedicamentuluiBrinza Irina-CarmenNoch keine Bewertungen
- Glucide - Curs 1, 2, 3, 4Dokument55 SeitenGlucide - Curs 1, 2, 3, 4Vasilica BoghiuNoch keine Bewertungen
- Transportul Prin Membrana PlasmaticaDokument13 SeitenTransportul Prin Membrana PlasmaticaFlorin DumitracheNoch keine Bewertungen
- Caractere Generale Ale Virusurilor Si Ale BacteriilorDokument29 SeitenCaractere Generale Ale Virusurilor Si Ale BacteriilorTeodor MindreanuNoch keine Bewertungen
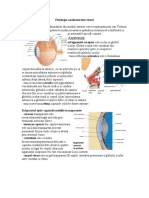
- Fiziologia Analizatorului VizualDokument2 SeitenFiziologia Analizatorului VizualPopa CarmenNoch keine Bewertungen
- Curs Nr. 10 Apoptoza Si NecrozaDokument67 SeitenCurs Nr. 10 Apoptoza Si NecrozaLarisa LașcuNoch keine Bewertungen
- Curs - GeneticăDokument24 SeitenCurs - GeneticăPaula UrsuNoch keine Bewertungen
- Supliment Enzime de RestrictieDokument2 SeitenSupliment Enzime de RestrictieCodrin MirceaNoch keine Bewertungen
- CELULADokument18 SeitenCELULAValentina Budurca100% (1)
- 1.2 Clasificarea Virusurilor În FamiliiDokument6 Seiten1.2 Clasificarea Virusurilor În FamiliiAna-Maria BarbuNoch keine Bewertungen
- DIABETDokument34 SeitenDIABETMarcu SanzianaNoch keine Bewertungen
- Prezentare Metabolismul GlucidelorDokument18 SeitenPrezentare Metabolismul GlucidelorMarina Denisa Ilina100% (1)
- Tabel Hormoni FiziologieDokument4 SeitenTabel Hormoni FiziologieMihai Hondru100% (1)
- Renal 1Dokument44 SeitenRenal 1Marcela Bucică100% (1)
- LP 10 - Histochimie +Dokument43 SeitenLP 10 - Histochimie +Andreea Maria BanicaNoch keine Bewertungen
- Subiecte Exam - An II - Semi Rezolvate.Dokument80 SeitenSubiecte Exam - An II - Semi Rezolvate.MonicaElenaNoch keine Bewertungen
- Curs 9Dokument51 SeitenCurs 9andrei100% (2)
- SangeleDokument11 SeitenSangelePopoiu GeorgeNoch keine Bewertungen
- Notiuni de Genetica UmanaDokument4 SeitenNotiuni de Genetica UmanaRemy DincaNoch keine Bewertungen
- Clostridiile LPDokument8 SeitenClostridiile LPCornelia HristiaNoch keine Bewertungen
- Absorbția IntestinalăDokument21 SeitenAbsorbția IntestinalăMarius-Sorin CionteaNoch keine Bewertungen
- Structura Analiza Şi Localizarea GenelorDokument48 SeitenStructura Analiza Şi Localizarea GenelorautorulaneiNoch keine Bewertungen
- FBAP - Acizii Nucleici Si Rolul Lor GeneticDokument10 SeitenFBAP - Acizii Nucleici Si Rolul Lor GeneticStefan Visan100% (1)
- Analiza CromozomilorDokument93 SeitenAnaliza Cromozomilorromanas25100% (1)
- Test Embriologie PDFDokument1 SeiteTest Embriologie PDFCrissy CrysNoch keine Bewertungen
- Curs Amg Singele Si T.muscularDokument53 SeitenCurs Amg Singele Si T.muscularAdina VioNoch keine Bewertungen
- SangeleDokument3 SeitenSangeleely_ely3395Noch keine Bewertungen
- 4'. Endocardite. Miocardite. PericarditeDokument55 Seiten4'. Endocardite. Miocardite. PericarditeTomoni StefanNoch keine Bewertungen
- Testul PCRDokument9 SeitenTestul PCRIulia BrindescuNoch keine Bewertungen
- Medicina SomnuluiDokument74 SeitenMedicina SomnuluiFuioaga Codrin-PaulNoch keine Bewertungen
- b.AciziiNucleici 2018Dokument59 Seitenb.AciziiNucleici 2018Dragos PrisacaruNoch keine Bewertungen
- Boli Mitocondriale REFERATDokument9 SeitenBoli Mitocondriale REFERATNdr PrlNoch keine Bewertungen
- Histologie 1 Curs 12) Tesutul NervosDokument7 SeitenHistologie 1 Curs 12) Tesutul NervosMariaMaryaNoch keine Bewertungen
- PoliomielitaDokument13 SeitenPoliomielitaVlada CebanNoch keine Bewertungen
- OrganiteDokument5 SeitenOrganiteLazy TurtleNoch keine Bewertungen
- Acizii Nucleici ADN Și ARNDokument8 SeitenAcizii Nucleici ADN Și ARNiulia bekeNoch keine Bewertungen
- Parazitologie MedicalaDokument46 SeitenParazitologie MedicalaAntonin Bogdan MitroaicaNoch keine Bewertungen
- Reticul EndoplasmaticDokument29 SeitenReticul EndoplasmaticbeibyloveNoch keine Bewertungen
- Metabolism LipidicDokument51 SeitenMetabolism LipidicCioltan AlexandraNoch keine Bewertungen
- Caile BiliareDokument11 SeitenCaile BiliareBulbasa BogdanNoch keine Bewertungen
- Ciuperci Paracetamol NitritiDokument63 SeitenCiuperci Paracetamol NitritiCami ManailaNoch keine Bewertungen
- Rezumat MicroDokument35 SeitenRezumat Microremus_stoica23Noch keine Bewertungen
- Curs NR 7 HormoniiDokument9 SeitenCurs NR 7 HormoniiCCCAXANoch keine Bewertungen
- Anatomie Alcatuire Corp UmanDokument7 SeitenAnatomie Alcatuire Corp Umanblume_weiss7651Noch keine Bewertungen
- Singe FiziologieDokument73 SeitenSinge FiziologieAnonymous ck0aGAqZl100% (1)
- Acizii Nucleici.Dokument42 SeitenAcizii Nucleici.Eugenia RusuNoch keine Bewertungen
- BiologieDokument9 SeitenBiologieStaver NataliaNoch keine Bewertungen
- Acizii NucleiciDokument63 SeitenAcizii Nucleiciana2586Noch keine Bewertungen
- Acizii NucleiciDokument8 SeitenAcizii NucleiciMarinela BurtescuNoch keine Bewertungen
- Acizii Nucleici 2Dokument146 SeitenAcizii Nucleici 2gvmeareason100% (1)
- ADNDokument47 SeitenADNCpt.WnchstrNoch keine Bewertungen
- Cap - 3 Cromozomul La Organisme Pro - Si EucarioteDokument13 SeitenCap - 3 Cromozomul La Organisme Pro - Si Eucariotecreatza20Noch keine Bewertungen
- Cap3-2 Enzime de RestrictieDokument8 SeitenCap3-2 Enzime de Restrictiecreatza20100% (1)
- Genetica Microorganismelor Şi Inginerie Genetică Microbiană NoteDokument19 SeitenGenetica Microorganismelor Şi Inginerie Genetică Microbiană Notecreatza20100% (4)
- Genetica Microorganismelor Şi Inginerie Genetică Microbiană NoteDokument13 SeitenGenetica Microorganismelor Şi Inginerie Genetică Microbiană Notecreatza20Noch keine Bewertungen
- Cap1 3 3 - TransductiaDokument4 SeitenCap1 3 3 - TransductiaAlina CatrangiuNoch keine Bewertungen
- Cap2-1 Tehnici Genetica Bacteriana ExtraDokument8 SeitenCap2-1 Tehnici Genetica Bacteriana Extracreatza20Noch keine Bewertungen
- Cap1-3-2 Transform Area Bacteriana ExtraDokument3 SeitenCap1-3-2 Transform Area Bacteriana Extracreatza20Noch keine Bewertungen
- Cap1-2-2 Plasmide Replicare ReglajDokument6 SeitenCap1-2-2 Plasmide Replicare ReglajLarisa-DNoch keine Bewertungen
- Cap1-1 - Cromozom BacterianDokument7 SeitenCap1-1 - Cromozom BacterianCristina AdamNoch keine Bewertungen
- Cap - 4 Functiile Materialului GeneticDokument20 SeitenCap - 4 Functiile Materialului Geneticcreatza20Noch keine Bewertungen
- Cap1-2-1 Plasmide Bacteriene STR Clasif ExtraDokument3 SeitenCap1-2-1 Plasmide Bacteriene STR Clasif Extracreatza20Noch keine Bewertungen
- Protocol de Explorare CardioDokument28 SeitenProtocol de Explorare Cardiocreatza20Noch keine Bewertungen
- A Functiei Excretorii+DigestivDokument32 SeitenA Functiei Excretorii+Digestivcreatza20Noch keine Bewertungen
- Bilant Functional Central+EndocrinDokument35 SeitenBilant Functional Central+Endocrincreatza20Noch keine Bewertungen