Das könnte Ihnen auch gefallen
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- LittorinidaeDokument358 SeitenLittorinidaeSyarif Prasetyo AdyutaNoch keine Bewertungen
- Complete Admin Law OutlineDokument135 SeitenComplete Admin Law Outlinemarlena100% (1)
- Respiratory Examination OSCE GuideDokument33 SeitenRespiratory Examination OSCE GuideBasmah 7Noch keine Bewertungen
- Capgras SyndromeDokument4 SeitenCapgras Syndromeapi-459379591Noch keine Bewertungen
- Faringitis StreptococcusDokument16 SeitenFaringitis StreptococcusnurNoch keine Bewertungen
- Final Box Design Prog.Dokument61 SeitenFinal Box Design Prog.afifa kausarNoch keine Bewertungen
- Invitation 2023Dokument10 SeitenInvitation 2023Joanna Marie Cruz FelipeNoch keine Bewertungen
- Accomplishment Report: For Service Credits (Availment/claim) Election May 2019Dokument1 SeiteAccomplishment Report: For Service Credits (Availment/claim) Election May 2019Glazy Kim Seco - JorquiaNoch keine Bewertungen
- RWE Algebra 12 ProbStat Discrete Math Trigo Geom 2017 DVO PDFDokument4 SeitenRWE Algebra 12 ProbStat Discrete Math Trigo Geom 2017 DVO PDFハンター ジェイソンNoch keine Bewertungen
- Contract of PledgeDokument4 SeitenContract of Pledgeshreya patilNoch keine Bewertungen
- Strategic Risk ManagementDokument46 SeitenStrategic Risk ManagementNuman Rox100% (1)
- Teuku Tahlil Prosiding38491Dokument30 SeitenTeuku Tahlil Prosiding38491unosa unounoNoch keine Bewertungen
- DigoxinDokument18 SeitenDigoxinApril Mergelle LapuzNoch keine Bewertungen
- HG G5 Q1 Mod1 RTP PDFDokument11 SeitenHG G5 Q1 Mod1 RTP PDFKimberly Abilon-Carlos100% (1)
- India: SupplyDokument6 SeitenIndia: SupplyHarish NathanNoch keine Bewertungen
- CLASS 12 PracticalDokument10 SeitenCLASS 12 PracticalWORLD HISTORYNoch keine Bewertungen
- Al-Tawheed According To The SahabahDokument16 SeitenAl-Tawheed According To The SahabahShadab AnjumNoch keine Bewertungen
- Operations Management (Scheduling) PDFDokument4 SeitenOperations Management (Scheduling) PDFVijay Singh ThakurNoch keine Bewertungen
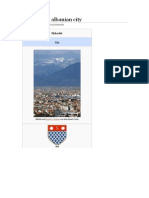
- Shkodër An Albanian CityDokument16 SeitenShkodër An Albanian CityXINKIANGNoch keine Bewertungen
- Popular Music Analysis and MusicologyDokument15 SeitenPopular Music Analysis and MusicologyAlexMartínVidal100% (3)
- Global Marketing & R&D CH 15Dokument16 SeitenGlobal Marketing & R&D CH 15Quazi Aritra ReyanNoch keine Bewertungen
- WPhO (Singapore) - World Physics Olympiad (WPhO) - 2011Dokument20 SeitenWPhO (Singapore) - World Physics Olympiad (WPhO) - 2011GXGGXG50% (2)
- ASME-Y14.5.1M 1994 Mathematical Definition of Dimensioning and Tolerancing Principles PDFDokument89 SeitenASME-Y14.5.1M 1994 Mathematical Definition of Dimensioning and Tolerancing Principles PDFwulfgang66Noch keine Bewertungen
- Summar Training Report HRTC TRAINING REPORTDokument43 SeitenSummar Training Report HRTC TRAINING REPORTPankaj ChauhanNoch keine Bewertungen
- Faith-Based Organisational Development (OD) With Churches in MalawiDokument10 SeitenFaith-Based Organisational Development (OD) With Churches in MalawiTransbugoyNoch keine Bewertungen
- MSDS PetrolDokument13 SeitenMSDS PetrolazlanNoch keine Bewertungen
- Level 5 - LFH 6-10 SepDokument14 SeitenLevel 5 - LFH 6-10 SepJanna GunioNoch keine Bewertungen
- Marcelo H Del PilarDokument8 SeitenMarcelo H Del PilarLee Antonino AtienzaNoch keine Bewertungen
- CopacabanaDokument2 SeitenCopacabanaNereus Sanaani CAñeda Jr.Noch keine Bewertungen
- Workshop 02.1: Restart Controls: ANSYS Mechanical Basic Structural NonlinearitiesDokument16 SeitenWorkshop 02.1: Restart Controls: ANSYS Mechanical Basic Structural NonlinearitiesSahil Jawa100% (1)