Das könnte Ihnen auch gefallen
- PRACTICA N 01 Determinacion de AzucaresDokument4 SeitenPRACTICA N 01 Determinacion de AzucaresJoiss ChirinosNoch keine Bewertungen
- Guia de Practica Morfofisiologia Normal IIDokument107 SeitenGuia de Practica Morfofisiologia Normal IIMRGM MUSICNoch keine Bewertungen
- Erupción Pasiva y ActivaDokument30 SeitenErupción Pasiva y ActivaSergio Llufire Mendoza100% (2)
- PhysioEx Ejercicio 10 Actividad 1Dokument3 SeitenPhysioEx Ejercicio 10 Actividad 1Carmen Pareja BetancourtNoch keine Bewertungen
- Presentación de Caso Clínico de Odontopediatría - MaylinDokument40 SeitenPresentación de Caso Clínico de Odontopediatría - MaylinSergio Llufire MendozaNoch keine Bewertungen
- PRACTICA 24. Determinación de Ácido Úrico PDFDokument3 SeitenPRACTICA 24. Determinación de Ácido Úrico PDFGaby Lisbeth Llh50% (4)
- PhysioEx Ejercicio 10 Actividad 3Dokument3 SeitenPhysioEx Ejercicio 10 Actividad 3Carmen Pareja Betancourt100% (1)
- Práctica 11-Digestión de CarbohidrátosDokument12 SeitenPráctica 11-Digestión de CarbohidrátosKEVIN JUNIOR RIVERA VASQUEZ100% (1)
- Guía de Elaboración Del Informe de Laboratorio y Prácticas de LaboratorioDokument36 SeitenGuía de Elaboración Del Informe de Laboratorio y Prácticas de LaboratorioKarleydis BarriosNoch keine Bewertungen
- Digestión Enzimática Del Almidón Bioquimica FinalDokument7 SeitenDigestión Enzimática Del Almidón Bioquimica FinalRoseline Galiano43% (7)
- Ortodoncia PrequirugicaDokument48 SeitenOrtodoncia PrequirugicaSergio Llufire MendozaNoch keine Bewertungen
- Sintesis de Acidos GrasosDokument22 SeitenSintesis de Acidos GrasosCITLALLI MU�OZ ANDRADENoch keine Bewertungen
- Práctica de Shock Espinal para Estudiar Los Reflejos en El SapoDokument11 SeitenPráctica de Shock Espinal para Estudiar Los Reflejos en El SapoTatiana Casas100% (3)
- Monografí Neoplasias Benignas en La Cavidad OralDokument32 SeitenMonografí Neoplasias Benignas en La Cavidad OralMichaelAnthonyWillyamDamiánLinaresNoch keine Bewertungen
- Copia de PRACTICA SHOCK ESPINAL o REFLEJ en SAPO Reflejos Osteo Tendinosos en El HombreUltimo MDokument19 SeitenCopia de PRACTICA SHOCK ESPINAL o REFLEJ en SAPO Reflejos Osteo Tendinosos en El HombreUltimo MMeli Baltazar T50% (6)
- Factores Que Intervienen en La Contracción MuscularDokument3 SeitenFactores Que Intervienen en La Contracción MuscularJuan Norris AldacoNoch keine Bewertungen
- Informe de Sapo Espinal Semana 8Dokument5 SeitenInforme de Sapo Espinal Semana 8MELANY RUBY HUANQUI CCALLO100% (1)
- 1 Informe de PracticaDokument7 Seiten1 Informe de PracticaPaul RuizNoch keine Bewertungen
- Práctica de La Amilasa SalivalDokument2 SeitenPráctica de La Amilasa SalivalBeatriz Martínez Ruiz100% (1)
- Depuración Renal, Aplicación de La Depuración Renal en La ClínicaDokument28 SeitenDepuración Renal, Aplicación de La Depuración Renal en La ClínicaVictorManuelAllendeQuispe100% (1)
- Espacio PelvisubperitonealDokument4 SeitenEspacio PelvisubperitonealYalitza Álvarez BarazarteNoch keine Bewertungen
- Concentracion y Dilucion UrinariaDokument2 SeitenConcentracion y Dilucion UrinariaAndrea La Rosa R50% (4)
- Mapa Articulaciones de La Cabeza Parte 1Dokument1 SeiteMapa Articulaciones de La Cabeza Parte 1Esli MedinaNoch keine Bewertungen
- AGA Disturbios Respiratorios PLUS MEDIC ADokument8 SeitenAGA Disturbios Respiratorios PLUS MEDIC AGretchen LavadoNoch keine Bewertungen
- ENZIMAS II TranscripcionDokument9 SeitenENZIMAS II Transcripcionapi-369665675% (4)
- Exploracion Fisica de Los Miembros InferioresDokument7 SeitenExploracion Fisica de Los Miembros InferioresLiz Mariam PortocarreroNoch keine Bewertungen
- 6) Embriologia SISTEMA RESPIRATORIO-1Dokument5 Seiten6) Embriologia SISTEMA RESPIRATORIO-1Jose Guillermo Mejia SaldanaNoch keine Bewertungen
- PhysioEx Ejercicio 3 Actividad 3 LAB 2Dokument4 SeitenPhysioEx Ejercicio 3 Actividad 3 LAB 2ESMILLSEN ANDREA QUISPE FLORESNoch keine Bewertungen
- Cardio. Preguntas de FisioDokument3 SeitenCardio. Preguntas de FisioVioleta Sequeiros VargasNoch keine Bewertungen
- Procedimento para La Determinacion de Calcio en SueroDokument2 SeitenProcedimento para La Determinacion de Calcio en SueroLaura Saavedra OlmosNoch keine Bewertungen
- Bulbo RaquideoDokument23 SeitenBulbo RaquideoRicardo PiguaveNoch keine Bewertungen
- AbdomenDokument22 SeitenAbdomenAnonymous 9nYdcc6LSNoch keine Bewertungen
- Pernineo Anterior FemeninoDokument36 SeitenPernineo Anterior Femeninojuanitaabigail50% (6)
- Informe de Medio InternoDokument14 SeitenInforme de Medio InternoAlmeyda Jefferson75% (4)
- Experimento Del Sistema Digestivo CULMINADODokument10 SeitenExperimento Del Sistema Digestivo CULMINADOMiguel Velez100% (1)
- PRACTICA 01. Determinación de Ácido ÚricoDokument3 SeitenPRACTICA 01. Determinación de Ácido ÚricosilviaNoch keine Bewertungen
- Informe Disección de SapoDokument6 SeitenInforme Disección de SapoJani AlarcónNoch keine Bewertungen
- Practica # 6 - Fosfatasa AlcalinaDokument7 SeitenPractica # 6 - Fosfatasa Alcalinarockakicha100% (1)
- Actividad 4 y 5 Lab Fisio RanaDokument3 SeitenActividad 4 y 5 Lab Fisio RanaCarlos Jair Herrera CamposNoch keine Bewertungen
- TSH NeonatalDokument5 SeitenTSH NeonatalLeandro NarváezNoch keine Bewertungen
- Regulación Hormonal de La Glucemia en Conejo 1Dokument4 SeitenRegulación Hormonal de La Glucemia en Conejo 1Denis R Gallardo Paredes100% (1)
- Practica 13Dokument2 SeitenPractica 13Andre HaroNoch keine Bewertungen
- Informe 2 - Determinación de La Concentración Sérica de Proteínas Totales y FraccionadasDokument10 SeitenInforme 2 - Determinación de La Concentración Sérica de Proteínas Totales y FraccionadasAlex MendivilNoch keine Bewertungen
- Informe Bioquímica Respiración TisularDokument7 SeitenInforme Bioquímica Respiración TisularEduardo Ramírez CapristánNoch keine Bewertungen
- VENOCLISIS11111Dokument4 SeitenVENOCLISIS11111Julio Cesar Valle InchausteNoch keine Bewertungen
- FISIOLOGIA, Acto Reflejo en SapoDokument9 SeitenFISIOLOGIA, Acto Reflejo en SapoSteven GeanpiereNoch keine Bewertungen
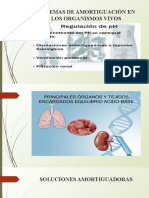
- Soluciones AmortiguadorasDokument19 SeitenSoluciones AmortiguadorasHade PhoccoNoch keine Bewertungen
- Carbohidratos Bioquimica USMPDokument26 SeitenCarbohidratos Bioquimica USMPBruno Avila Veneros100% (2)
- Ejercicio PhysioEx 3 Actividad 7Dokument4 SeitenEjercicio PhysioEx 3 Actividad 7Karla HuamaniNoch keine Bewertungen
- Pelvis EspinozaDokument131 SeitenPelvis EspinozaRenzo RiveraNoch keine Bewertungen
- Práctica #1 de Laboratorio Fisiología Neuromuscular y de Los Sistemas Sensoriales 2016Dokument6 SeitenPráctica #1 de Laboratorio Fisiología Neuromuscular y de Los Sistemas Sensoriales 2016Carlos Aquise LaricoNoch keine Bewertungen
- Síntesis, Transporte y Excreción de ColesterolDokument59 SeitenSíntesis, Transporte y Excreción de ColesterolIsabel100% (1)
- Histologia Tejido Muscular Cardiaco y LisoDokument18 SeitenHistologia Tejido Muscular Cardiaco y LisoYami StrikeNoch keine Bewertungen
- Resuelto PhysioEx Ejercicio 9 Actividad 4Dokument2 SeitenResuelto PhysioEx Ejercicio 9 Actividad 4Lucia Saavedra Lozada100% (1)
- Pràctica #5 Determinación de CalcioDokument11 SeitenPràctica #5 Determinación de CalcioNelly Estefany PechoNoch keine Bewertungen
- Trabajo 5.Dokument4 SeitenTrabajo 5.Bruno QuispeNoch keine Bewertungen
- Tinción de WrightDokument2 SeitenTinción de WrightPaola Ortega FrancoNoch keine Bewertungen
- Practica4 FisiologiaDokument3 SeitenPractica4 FisiologiaJuan Diego Cahuana VasquezNoch keine Bewertungen
- Practica 03 de Quimica ListaDokument5 SeitenPractica 03 de Quimica ListaLorena Izquierdo sabino100% (1)
- Parcial MorfofisiologiaDokument14 SeitenParcial MorfofisiologiaMerilynBridgetNoch keine Bewertungen
- Artículo Soluciones BufferDokument5 SeitenArtículo Soluciones BufferJMauricio21Noch keine Bewertungen
- Guia Fisiologia I PDFDokument151 SeitenGuia Fisiologia I PDFmadelyn aydeeNoch keine Bewertungen
- Sumacion Temporal y Espacial Informe DsssssssssssssssssssssssssssssssssssssDokument19 SeitenSumacion Temporal y Espacial Informe DsssssssssssssssssssssssssssssssssssssWalterQunoqYanceOwNoch keine Bewertungen
- Taller BiologiaDokument6 SeitenTaller BiologiaAnderson50% (6)
- Trabajo de BiologiaDokument9 SeitenTrabajo de BiologiaRodrigo Navarrete SotoNoch keine Bewertungen
- Prueba Validez Contenido-InstrumDokument5 SeitenPrueba Validez Contenido-InstrumSergio Llufire MendozaNoch keine Bewertungen
- Principios de TalladoDokument45 SeitenPrincipios de TalladoSergio Llufire MendozaNoch keine Bewertungen
- Plan de TratamientoDokument33 SeitenPlan de TratamientoSergio Llufire MendozaNoch keine Bewertungen
- Alargamiento Coronario Una Necesidad de Retencion Protesica Estetica y Anchura Biologica Revision BibliograficaDokument10 SeitenAlargamiento Coronario Una Necesidad de Retencion Protesica Estetica y Anchura Biologica Revision BibliograficaSergio Llufire MendozaNoch keine Bewertungen
- Ciclo CelularDokument25 SeitenCiclo CelularLaura MoraNoch keine Bewertungen
- Tema 3 Adaptacion CelularDokument24 SeitenTema 3 Adaptacion CelularMildred ChavezNoch keine Bewertungen
- La Triada EcológicaDokument14 SeitenLa Triada EcológicaAndrea MuñozNoch keine Bewertungen
- Clase #1 INTRODUCCION A LA MICROBIOLOGIADokument66 SeitenClase #1 INTRODUCCION A LA MICROBIOLOGIAAlfred MárquezNoch keine Bewertungen
- Reporte 1. Extracción de ADNDokument13 SeitenReporte 1. Extracción de ADNElizabethNoch keine Bewertungen
- Metabolism oDokument10 SeitenMetabolism oTitto MéndezNoch keine Bewertungen
- ADL MetabolismoDokument18 SeitenADL MetabolismoDiana ZavalaNoch keine Bewertungen
- GUIAS DE APRENDIZAJE #7 Oscar Garcia GRADO SEXTO Ciencias Naturales-ArtisticaDokument12 SeitenGUIAS DE APRENDIZAJE #7 Oscar Garcia GRADO SEXTO Ciencias Naturales-ArtisticaOscarAugustoGarciaAlvarezNoch keine Bewertungen
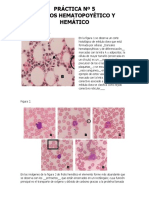
- Práctica 5. Tejido Hemático y Hematopoyético - EjerciciosDokument2 SeitenPráctica 5. Tejido Hemático y Hematopoyético - EjerciciosDaav L. GuarneriNoch keine Bewertungen
- Tarea Sobre La Redacción de Un Cuadro Sinóptico Sobre Las Semejanzas y Diferencias Entre ADN y ARN. Sergio Ramirez.Dokument3 SeitenTarea Sobre La Redacción de Un Cuadro Sinóptico Sobre Las Semejanzas y Diferencias Entre ADN y ARN. Sergio Ramirez.SERGIO DARWIN RAMIREZ LINARESNoch keine Bewertungen
- 23 Estres OxidativoDokument52 Seiten23 Estres OxidativoMiguel Chirinos AlarconNoch keine Bewertungen
- PENICILINAaaaaaaaaaaaaaaaaaaaaaaaaaaaDokument16 SeitenPENICILINAaaaaaaaaaaaaaaaaaaaaaaaaaaaGaby MorenoNoch keine Bewertungen
- Infografía - Ácidos NucleicosDokument13 SeitenInfografía - Ácidos NucleicosAlejandra SanchezNoch keine Bewertungen
- Tecnicas de Eliminacion de MicroorganismosDokument7 SeitenTecnicas de Eliminacion de Microorganismoscanva cuentaNoch keine Bewertungen
- Evolucion Perfil LipidicoDokument10 SeitenEvolucion Perfil LipidicoAlex JaimesNoch keine Bewertungen
- Rúbrica para Evaluar Un CartelDokument7 SeitenRúbrica para Evaluar Un CartelCarlos VillalobosNoch keine Bewertungen
- Trabajo Final Prevencion Del Cancer3Dokument15 SeitenTrabajo Final Prevencion Del Cancer3MoisesTorresSNoch keine Bewertungen
- Formato Informe FotosíntesisDokument10 SeitenFormato Informe FotosíntesisCamilo Andres Rico CortesNoch keine Bewertungen
- Degradacion de Los Aminoacidos (Bioqumica)Dokument3 SeitenDegradacion de Los Aminoacidos (Bioqumica)KAaTHERINnNoch keine Bewertungen
- 5to Trabajo PrácticoDokument9 Seiten5to Trabajo PrácticoIAMCAŁVØ GGNoch keine Bewertungen
- Glucotransportadores MapaDokument1 SeiteGlucotransportadores MapaViridiana Hernandez DiazNoch keine Bewertungen
- Resumen Origen de La Vida - ShapiroDokument3 SeitenResumen Origen de La Vida - ShapiroJair Rhodriwez100% (1)
- CloroplastosDokument5 SeitenCloroplastosIvonne PadillaNoch keine Bewertungen
- Efectos Adversos Del TARGADokument25 SeitenEfectos Adversos Del TARGAAnonymous Kv0sHqFNoch keine Bewertungen
- HUASCARANDokument95 SeitenHUASCARANJavier MolinaNoch keine Bewertungen
- Que Es El BHB - Buscar Con GoogleDokument2 SeitenQue Es El BHB - Buscar Con GoogleKachimira Da SilvaNoch keine Bewertungen
- Phaseolus Vulgaris IIDokument54 SeitenPhaseolus Vulgaris IIJonathan Harvey Poveda RicoNoch keine Bewertungen
- T Ucsg Pos Egm CP 8Dokument49 SeitenT Ucsg Pos Egm CP 8jorgeNoch keine Bewertungen