Das könnte Ihnen auch gefallen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (894)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (587)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (119)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2219)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Coesel Et Al, 2017 PDFDokument7 SeitenCoesel Et Al, 2017 PDFmilerakeNoch keine Bewertungen
- Ecological Distribution of Cyanophyceae in Lotic Ecosystems of São Paulo StateDokument10 SeitenEcological Distribution of Cyanophyceae in Lotic Ecosystems of São Paulo StatemilerakeNoch keine Bewertungen
- LIBRO Marine Phytoplankton Atlas of Kuwait's WatersDokument351 SeitenLIBRO Marine Phytoplankton Atlas of Kuwait's Watersjchuatson50% (2)
- 2011 Lib Rode Practic AsDokument121 Seiten2011 Lib Rode Practic AsmilerakeNoch keine Bewertungen
- Dinoflagelados de Australia PDFDokument55 SeitenDinoflagelados de Australia PDFmilerakeNoch keine Bewertungen
- Sistema Nervioso Del GrupoDokument9 SeitenSistema Nervioso Del GrupomilerakeNoch keine Bewertungen
- LIBRO Marine Phytoplankton Atlas of Kuwait's WatersDokument351 SeitenLIBRO Marine Phytoplankton Atlas of Kuwait's Watersjchuatson50% (2)
- Extension of Flood PulseDokument23 SeitenExtension of Flood PulsemilerakeNoch keine Bewertungen
- Phoronida TamañoDokument7 SeitenPhoronida TamañomilerakeNoch keine Bewertungen
- Marine Flora and Fauna.2Dokument11 SeitenMarine Flora and Fauna.2MissJalene ObradorNoch keine Bewertungen
- Algal Origin and DiversityDokument18 SeitenAlgal Origin and DiversityArunchand RayarothNoch keine Bewertungen
- Advances in Phycological StudiesDokument385 SeitenAdvances in Phycological StudiesItachi Mtnez50% (2)
- Uas 2020Dokument5 SeitenUas 2020fiqi Hidayat FamirNoch keine Bewertungen
- Phytoplankton Community Structure in The River-Influenced Continental Margin of The Northern Gulf of MexicoDokument17 SeitenPhytoplankton Community Structure in The River-Influenced Continental Margin of The Northern Gulf of MexicoZelfi IndrawatyNoch keine Bewertungen
- Studies On The Diatom Flora of North Maharashtra Region Genus - CaloneisDokument24 SeitenStudies On The Diatom Flora of North Maharashtra Region Genus - CaloneisESSENCE - International Journal for Environmental Rehabilitation and ConservaionNoch keine Bewertungen
- Description MicroscopeDokument4 SeitenDescription MicroscopeRanma SaotomeNoch keine Bewertungen
- Science Magazine 5693 2004-10-01Dokument107 SeitenScience Magazine 5693 2004-10-01WillimSmith100% (1)
- Ekologi Mikroalga - 2013Dokument36 SeitenEkologi Mikroalga - 2013ChoNoch keine Bewertungen
- 2023 - George Felfoldi - (Ebook - Health) - Another Look at Seaweed and Algae, 136 PagesDokument136 Seiten2023 - George Felfoldi - (Ebook - Health) - Another Look at Seaweed and Algae, 136 PagesGeorge FelfoldiNoch keine Bewertungen
- Marine Microfossils: A Guide to Microscopic Remains in Ocean SedimentsDokument9 SeitenMarine Microfossils: A Guide to Microscopic Remains in Ocean SedimentsHalima akterNoch keine Bewertungen
- Algae TestDokument3 SeitenAlgae TestSeika SaliNoch keine Bewertungen
- Earth Sciences Notes Csir-Ugc Net JRFDokument57 SeitenEarth Sciences Notes Csir-Ugc Net JRFmanas773Noch keine Bewertungen
- Stromatolites Forming in Acidic Hot-Spring WatersDokument26 SeitenStromatolites Forming in Acidic Hot-Spring Watersgigio marinoNoch keine Bewertungen
- Publication 12 28628 754Dokument12 SeitenPublication 12 28628 754Keny Ruslan Mowilos100% (1)
- Janse Van Vuuren 2006 Easy Identification of The Most Common Freshwater AlgaeDokument212 SeitenJanse Van Vuuren 2006 Easy Identification of The Most Common Freshwater Algaejulio.nunez2901100% (5)
- Comparing Three Schools of Thought on Algal ClassificationDokument9 SeitenComparing Three Schools of Thought on Algal ClassificationMichelNoch keine Bewertungen
- Domain Eukarya-YR10 - PART2 - TUTE2Dokument7 SeitenDomain Eukarya-YR10 - PART2 - TUTE2Sandalu Romeshika PereraNoch keine Bewertungen
- Heterokont Chloroplast EvolutionDokument22 SeitenHeterokont Chloroplast EvolutionsenorgustoNoch keine Bewertungen
- 3 Plankton Guide 2010revDokument22 Seiten3 Plankton Guide 2010revsolo baru075Noch keine Bewertungen
- Beyond Micromachining - The Potential of DiatomsDokument7 SeitenBeyond Micromachining - The Potential of DiatomsYunuhen BadilloNoch keine Bewertungen
- Classification of Algae Divided into 11 ClassesDokument6 SeitenClassification of Algae Divided into 11 ClassessankarNoch keine Bewertungen
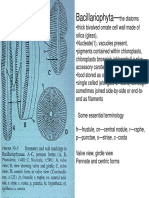
- Bacillariophyta—the Diatoms and Their Unique Silica Cell WallsDokument23 SeitenBacillariophyta—the Diatoms and Their Unique Silica Cell WallsSwetha RameshNoch keine Bewertungen
- Emerciano, Et Al. 2017. BIofloc (Generalidades) Plancton PDFDokument21 SeitenEmerciano, Et Al. 2017. BIofloc (Generalidades) Plancton PDFAlejandro Giraldo ObregónNoch keine Bewertungen
- International Paleolimnology Symposium 12th IPS 2012Dokument214 SeitenInternational Paleolimnology Symposium 12th IPS 2012vmsolartecNoch keine Bewertungen
- 5 - MicroalgaeDokument26 Seiten5 - MicroalgaeRUSSEL MAE CATIMBANGNoch keine Bewertungen
- Diatom Algae-Nature Is Helping UsDokument18 SeitenDiatom Algae-Nature Is Helping UsAndreas AbrahamNoch keine Bewertungen
- (Acta Palaeobotanica) On The Age of Fossil DiatomsDokument3 Seiten(Acta Palaeobotanica) On The Age of Fossil DiatomsArvind NegiNoch keine Bewertungen
- Illustrated River Phytoplankton PDFDokument68 SeitenIllustrated River Phytoplankton PDFMühåmmãd Îlhäm Zãrqøñý0% (1)
- Gracilaria Handbook Culture PDFDokument49 SeitenGracilaria Handbook Culture PDFFaizatulAsmaJaidinNoch keine Bewertungen