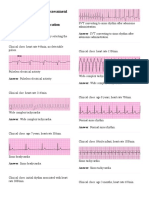
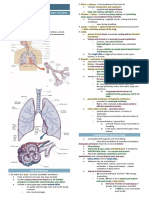
Das könnte Ihnen auch gefallen
- Psychiatry 101Dokument17 SeitenPsychiatry 101Vaso Talakvadze0% (1)
- The History of PsychiatryDokument12 SeitenThe History of PsychiatryMarkoff Chaney100% (1)
- Perinatology Clinics 2008, Vol.35, Issues 4, Neuroprotection in The NewbornDokument210 SeitenPerinatology Clinics 2008, Vol.35, Issues 4, Neuroprotection in The NewbornJhonny MarquezNoch keine Bewertungen
- MCU 2019 Blueprints Neurology 5th EditionDokument464 SeitenMCU 2019 Blueprints Neurology 5th EditionPermata GBKP Runggun Surabaya100% (2)
- Surgery OSCE QuestionsDokument27 SeitenSurgery OSCE QuestionsDineish MurugaiahNoch keine Bewertungen
- Diarrhea in ChildrenDokument42 SeitenDiarrhea in ChildrenIPNATC NEPALNoch keine Bewertungen
- Bone Infections - Acute OsteomyelitisDokument35 SeitenBone Infections - Acute Osteomyelitismeto40% (5)
- PALS Precourse Self 2023Dokument9 SeitenPALS Precourse Self 2023Airene SibleNoch keine Bewertungen
- CXCL12 PDFDokument17 SeitenCXCL12 PDFJean andre CanoNoch keine Bewertungen
- 4030 FullDokument10 Seiten4030 FullKartikeya SinghNoch keine Bewertungen
- Awab 134Dokument12 SeitenAwab 134yi1zhangNoch keine Bewertungen
- Regulation of Extracellular Superoxide Dismutase in Sertoli CellsDokument13 SeitenRegulation of Extracellular Superoxide Dismutase in Sertoli CellsHafiz MahruzzaNoch keine Bewertungen
- Specific Binding of RBC To EC Is Regulated by Nonadsorbing MacromoleculesDokument7 SeitenSpecific Binding of RBC To EC Is Regulated by Nonadsorbing MacromoleculesaguslistyNoch keine Bewertungen
- Phosphate Regulation of Vascular SmoothDokument9 SeitenPhosphate Regulation of Vascular Smoothbryan chanNoch keine Bewertungen
- Effect of PH On BMSCDokument8 SeitenEffect of PH On BMSCVishwarup GoswamiNoch keine Bewertungen
- CD11b ExpressionDokument4 SeitenCD11b ExpressionTisha Patricia OedoyNoch keine Bewertungen
- AKT/Protein Kinase B Regulation of BCL Family Members During Oxysterol-Induced ApoptosisDokument8 SeitenAKT/Protein Kinase B Regulation of BCL Family Members During Oxysterol-Induced ApoptosisNareshNoch keine Bewertungen
- Michael Kyba Et Al - Enhanced Hematopoietic Differentiation of Embryonic Stem Cells Conditionally Expressing Stat5Dokument7 SeitenMichael Kyba Et Al - Enhanced Hematopoietic Differentiation of Embryonic Stem Cells Conditionally Expressing Stat5Gmewop30mNoch keine Bewertungen
- Cai Et Al. - 2012 - The Heme Oxygenase 1 Inducer (CoPP) Protects Human Cardiac Stem Cells Against Apoptosis Through Activation of The ExDokument17 SeitenCai Et Al. - 2012 - The Heme Oxygenase 1 Inducer (CoPP) Protects Human Cardiac Stem Cells Against Apoptosis Through Activation of The Exscribd mendeleyNoch keine Bewertungen
- FA - Folic Acid-Mediated Fibrosis Is Driven by C5a Receptor 1-Mediated Activation of Kidney Myeloid CellsDokument14 SeitenFA - Folic Acid-Mediated Fibrosis Is Driven by C5a Receptor 1-Mediated Activation of Kidney Myeloid CellsCher IshNoch keine Bewertungen
- PL00000249 PDFDokument8 SeitenPL00000249 PDFdavid stNoch keine Bewertungen
- Jurnal LosartanDokument4 SeitenJurnal Losartanaditya_permana77Noch keine Bewertungen
- Baj Krzyworzeka (2002)Dokument10 SeitenBaj Krzyworzeka (2002)Sitara AkbarNoch keine Bewertungen
- Overexpression of Hematopoietically Expressed Homeoprotein Induces Nonapoptotic Cell Death in Mouse Prechondrogenic ATDC5 CellsDokument7 SeitenOverexpression of Hematopoietically Expressed Homeoprotein Induces Nonapoptotic Cell Death in Mouse Prechondrogenic ATDC5 CellsCamilo CabreraNoch keine Bewertungen
- Phil. Trans. R. Soc. B 2011 Unternaehrer 2274 85Dokument13 SeitenPhil. Trans. R. Soc. B 2011 Unternaehrer 2274 85Jo-Anne M. VentiganNoch keine Bewertungen
- Rapid Publication: Sickle CellDokument5 SeitenRapid Publication: Sickle CellVibhav SinghNoch keine Bewertungen
- Intravenous Infusion of Nerve Growth Factor-Secreting Monocytes Supports The Survival of Cholinergic Neurons in The Nucleus Basalis of Meynert in Hypercholesterolemia Brown-Norway RatsDokument9 SeitenIntravenous Infusion of Nerve Growth Factor-Secreting Monocytes Supports The Survival of Cholinergic Neurons in The Nucleus Basalis of Meynert in Hypercholesterolemia Brown-Norway Ratsdimitri pizarnikNoch keine Bewertungen
- Production of Functional Glucagon-Secreting From Human Embryonic Stem CellsDokument9 SeitenProduction of Functional Glucagon-Secreting From Human Embryonic Stem CellsNyna KawlesNoch keine Bewertungen
- Reactive Oxygen and Nitrogen Species in Sepsis-Induced Hepatic Microvascular DysfunctionDokument10 SeitenReactive Oxygen and Nitrogen Species in Sepsis-Induced Hepatic Microvascular DysfunctionGiosué Reyes HernandezNoch keine Bewertungen
- 3613 FullDokument10 Seiten3613 FullTisekNoch keine Bewertungen
- Potent Vasoconstrictor Kisspeptin-10 Induces Atherosclerotic Plaque Progression and Instability: Reversal by Its Receptor GPR54 AntagonistDokument20 SeitenPotent Vasoconstrictor Kisspeptin-10 Induces Atherosclerotic Plaque Progression and Instability: Reversal by Its Receptor GPR54 AntagonistÁlex GómezNoch keine Bewertungen
- Liv 12252Dokument9 SeitenLiv 12252Loli HunterNoch keine Bewertungen
- Atherosclerosis: Evidence For Impairment of Resolution of Vascular Inflammation Governed by Specific Lipid MediatorsDokument12 SeitenAtherosclerosis: Evidence For Impairment of Resolution of Vascular Inflammation Governed by Specific Lipid MediatorsMaria AnghelacheNoch keine Bewertungen
- Citosol (Thiamylal Sodium) Triggers Apoptosis and Affects Gene Expressions of Murine Leukemia RAW 264.7 Cells - RS-C Wu, C-S Yu, 2Dokument1 SeiteCitosol (Thiamylal Sodium) Triggers Apoptosis and Affects Gene Expressions of Murine Leukemia RAW 264.7 Cells - RS-C Wu, C-S Yu, 2Alondra MaldonadoNoch keine Bewertungen
- Anucleate Platelets Generate Progeny 1Dokument34 SeitenAnucleate Platelets Generate Progeny 1Alejandro BarretoNoch keine Bewertungen
- Microscopy Res Technique - 2001 - Southwood - Molecular Pathways of Oligodendrocyte Apoptosis Revealed by Mutations inDokument9 SeitenMicroscopy Res Technique - 2001 - Southwood - Molecular Pathways of Oligodendrocyte Apoptosis Revealed by Mutations inAnglia LopesNoch keine Bewertungen
- Endotelial Pericites - Cui2006Dokument10 SeitenEndotelial Pericites - Cui2006Lilo FrancoisNoch keine Bewertungen
- Chromosomal Aberrations Induced by 5-Azacytidine Combined With VP-16 (Etoposide) in CHO-K1 and XRS-5 Cell LinesDokument16 SeitenChromosomal Aberrations Induced by 5-Azacytidine Combined With VP-16 (Etoposide) in CHO-K1 and XRS-5 Cell LinesAry AguiarNoch keine Bewertungen
- 1984 Schlissel Virus-Transformed Pre-B Cells Show Ordered Activation But Not Inactivation of Immunoglobulin Gene Rearrangement and TranscriptionDokument10 Seiten1984 Schlissel Virus-Transformed Pre-B Cells Show Ordered Activation But Not Inactivation of Immunoglobulin Gene Rearrangement and TranscriptionKenneth YuNoch keine Bewertungen
- CationicGradientReversalandCytoskeleton IndependentDokument11 SeitenCationicGradientReversalandCytoskeleton Independentthymine613Noch keine Bewertungen
- Agrin en HematopoiesisDokument11 SeitenAgrin en HematopoiesisMarcos CollazoNoch keine Bewertungen
- Loss of PKCD Results in Characteristics of Sjo Gren'S Syndrome Including Salivary Gland DysfunctionDokument10 SeitenLoss of PKCD Results in Characteristics of Sjo Gren'S Syndrome Including Salivary Gland DysfunctiondrjatinNoch keine Bewertungen
- PLOS One-6-E21250-2011Dokument16 SeitenPLOS One-6-E21250-2011Sandi Head McAdamsNoch keine Bewertungen
- c1 PDFDokument9 Seitenc1 PDFoaifoiweuNoch keine Bewertungen
- Increased Islet Apoptosis in Pdx1 MiceDokument14 SeitenIncreased Islet Apoptosis in Pdx1 Miceroh009Noch keine Bewertungen
- Leukemia Stem Cells: Eike C. Buss and Anthony D. HoDokument9 SeitenLeukemia Stem Cells: Eike C. Buss and Anthony D. HoPedro AzevedoNoch keine Bewertungen
- TaylorDokument11 SeitenTaylorDawrul IslamNoch keine Bewertungen
- Stem Cell: The Spleen Contributes Stem Cells To Peripheral Blood Stem Cell TransplantsDokument4 SeitenStem Cell: The Spleen Contributes Stem Cells To Peripheral Blood Stem Cell TransplantsAnuj MittalNoch keine Bewertungen
- J Cell Sci-1993-Perou-99-107Dokument9 SeitenJ Cell Sci-1993-Perou-99-107glodovichiNoch keine Bewertungen
- Canine Malignant Hemangiosarcoma As A Model of Primitive Angiogenic EndotheliumDokument11 SeitenCanine Malignant Hemangiosarcoma As A Model of Primitive Angiogenic Endotheliumfsuarez113Noch keine Bewertungen
- Translational Oncology: MD Masud Alam, Ryusho Kariya, Piyanard Boonnate, Azusa Kawaguchi, Seiji OkadaDokument10 SeitenTranslational Oncology: MD Masud Alam, Ryusho Kariya, Piyanard Boonnate, Azusa Kawaguchi, Seiji OkadaAthanasios S. ArampatzisNoch keine Bewertungen
- Nelson 2015Dokument8 SeitenNelson 2015Thanasis PapatheodorouNoch keine Bewertungen
- Development of Anti-Peptide Polyclonal Antibodies Raised Against Immunogenic Sequences of Protein Encoded by Fj194940.1 GeneDokument9 SeitenDevelopment of Anti-Peptide Polyclonal Antibodies Raised Against Immunogenic Sequences of Protein Encoded by Fj194940.1 GeneijsidonlineinfoNoch keine Bewertungen
- 2009 Kang - Insulin Secreting Cells From Human Eyelid Derived Stem Cells Alleviate Type I Diabetes in Immunocompetent MiceDokument10 Seiten2009 Kang - Insulin Secreting Cells From Human Eyelid Derived Stem Cells Alleviate Type I Diabetes in Immunocompetent MiceNyna KawlesNoch keine Bewertungen
- Biochemical and Biophysical Research CommunicationsDokument7 SeitenBiochemical and Biophysical Research CommunicationsGiulia AndreeaNoch keine Bewertungen
- DLK1 As A Potential Target Against Cancer Stem/Progenitor Cells of Hepatocellular CarcinomaDokument10 SeitenDLK1 As A Potential Target Against Cancer Stem/Progenitor Cells of Hepatocellular CarcinomaHadrien DemagnyNoch keine Bewertungen
- Origins of Circulating Endothelial Cells and Endothelial Outgrowth From BloodDokument7 SeitenOrigins of Circulating Endothelial Cells and Endothelial Outgrowth From BloodsadhanaaNoch keine Bewertungen
- Efficient Stage-Specific Differentiation of Human Pluripotent Stem Cells Toward Retinal Photoreceptor Cells - Mellough 2012Dokument15 SeitenEfficient Stage-Specific Differentiation of Human Pluripotent Stem Cells Toward Retinal Photoreceptor Cells - Mellough 2012Rebeca ChávezNoch keine Bewertungen
- Gangliosides Expressed On Breast Cancer Cells Are E-Selectin LigandsDokument7 SeitenGangliosides Expressed On Breast Cancer Cells Are E-Selectin LigandsAnchalia ChandrakumaranNoch keine Bewertungen
- 2022 Balzer Fibrotic Kidney RegenerationDokument18 Seiten2022 Balzer Fibrotic Kidney RegenerationHeliosjafNoch keine Bewertungen
- Anemia Falciforme George2010 LeidoDokument5 SeitenAnemia Falciforme George2010 LeidoLaura CorreaNoch keine Bewertungen
- Tumor Necrosis Factor A Stimulates Human Clara Cell Secretory Protein Production by Human Airway Epithelial Cells.1998Dokument7 SeitenTumor Necrosis Factor A Stimulates Human Clara Cell Secretory Protein Production by Human Airway Epithelial Cells.1998Fina Herlinda NurNoch keine Bewertungen
- Nichos HematopoyeticosDokument10 SeitenNichos Hematopoyeticosmarcel_dcNoch keine Bewertungen
- NIH Public Access: Author ManuscriptDokument19 SeitenNIH Public Access: Author Manuscriptaldo123456789Noch keine Bewertungen
- Brief Definitive ReportDokument6 SeitenBrief Definitive ReportenilNoch keine Bewertungen
- Chloremphenicol Leukemia LinkedDokument17 SeitenChloremphenicol Leukemia LinkedpadbidriNoch keine Bewertungen
- Haem 2007 cxcr4 MSC HomingDokument8 SeitenHaem 2007 cxcr4 MSC HomingoaifoiweuNoch keine Bewertungen
- To Rejuvenate With The Blood Plasma Of Young PeopleVon EverandTo Rejuvenate With The Blood Plasma Of Young PeopleNoch keine Bewertungen
- Painful Hashimoto's Thyroiditis: EditorialDokument2 SeitenPainful Hashimoto's Thyroiditis: EditorialNovia MentariNoch keine Bewertungen
- 6Dokument23 Seiten6Novia MentariNoch keine Bewertungen
- What Is Hashimoto's Disease?Dokument6 SeitenWhat Is Hashimoto's Disease?Novia MentariNoch keine Bewertungen
- 2Dokument21 Seiten2Novia MentariNoch keine Bewertungen
- Reproductive Health (English)Dokument81 SeitenReproductive Health (English)proluvieslacusNoch keine Bewertungen
- E1.1. Reproduction and FertilityDokument47 SeitenE1.1. Reproduction and FertilityNovia MentariNoch keine Bewertungen
- Hunter and Arlin B. Blood Gordon G. Power, Shannon L. Bragg, Bryan T. Oshiro, Andre Dejam, Christian JDokument8 SeitenHunter and Arlin B. Blood Gordon G. Power, Shannon L. Bragg, Bryan T. Oshiro, Andre Dejam, Christian JNovia MentariNoch keine Bewertungen
- UnairDokument1 SeiteUnairNovia MentariNoch keine Bewertungen
- 2011-2012 Annual Report Department of Pediatrics Saint Louis University School of MedicineDokument131 Seiten2011-2012 Annual Report Department of Pediatrics Saint Louis University School of Medicineslusompeds0% (1)
- Fdar Cabg 3nesurgicalDokument5 SeitenFdar Cabg 3nesurgicalMary Margareth GonzalesNoch keine Bewertungen
- Deyo 1992Dokument6 SeitenDeyo 1992Wwwanand111Noch keine Bewertungen
- LVADDokument23 SeitenLVADVashish RamrechaNoch keine Bewertungen
- Approach To Delay PubertyDokument33 SeitenApproach To Delay PubertyRaja Iqbal Mulya HarahapNoch keine Bewertungen
- Jemds Title PDokument1 SeiteJemds Title PKriti KumariNoch keine Bewertungen
- NCP - Activity IntoleranceDokument5 SeitenNCP - Activity IntolerancePrincess Alane MorenoNoch keine Bewertungen
- Elc501 - Group Portfolio ForumDokument11 SeitenElc501 - Group Portfolio ForumAhmad SyamiNoch keine Bewertungen
- The LA Classification of GERDDokument2 SeitenThe LA Classification of GERDelisa100% (1)
- Procedure: Inserting A Nasogastric/ Nasointestinal Tube: BUENO, Jessa Mae D 3-8 February 28, 2024Dokument15 SeitenProcedure: Inserting A Nasogastric/ Nasointestinal Tube: BUENO, Jessa Mae D 3-8 February 28, 2024Jessa BuenoNoch keine Bewertungen
- Covid-19: A Recommendation To Global Health PolicyDokument5 SeitenCovid-19: A Recommendation To Global Health PolicyDr. Nur-E-Alam Rasel50% (2)
- Oxygenation Structure of The Respiratory System: Pulmonary VentilationDokument40 SeitenOxygenation Structure of The Respiratory System: Pulmonary VentilationCaitlin G.Noch keine Bewertungen
- 10 1 1 115Dokument151 Seiten10 1 1 115Angela TorrenteNoch keine Bewertungen
- Swine Quiz Bowl QuestionsDokument6 SeitenSwine Quiz Bowl QuestionsNezuko CutieeeNoch keine Bewertungen
- Dizziness HXDokument4 SeitenDizziness HXbadmanNoch keine Bewertungen
- Drug InteractionDokument151 SeitenDrug Interactionapi-3724213100% (1)
- Dysbiosis of The Human Oral Microbiome During The Menstrual Cycle and Vulnerability To The External Exposures of Smoking and Dietary SugarDokument14 SeitenDysbiosis of The Human Oral Microbiome During The Menstrual Cycle and Vulnerability To The External Exposures of Smoking and Dietary SugarPaloma C. PeñaNoch keine Bewertungen
- Peritonitis Update On Pathophysiology, Clinical Manifestations, and PDFDokument11 SeitenPeritonitis Update On Pathophysiology, Clinical Manifestations, and PDFAnonymous InJS6aYZ100% (1)
- Nso EgdDokument3 SeitenNso Egdtry dokkNoch keine Bewertungen
- Nhis Drug Price List Final (2013 Ed)Dokument32 SeitenNhis Drug Price List Final (2013 Ed)holuwadamilare935Noch keine Bewertungen
- WK 4 Respiratory 2023Dokument38 SeitenWK 4 Respiratory 2023Basmala HebaNoch keine Bewertungen
- PROGRAMME ABSTRACT BOOK IACS ES 4 7 Oct 2023 Timisoara Romania - CORRDokument164 SeitenPROGRAMME ABSTRACT BOOK IACS ES 4 7 Oct 2023 Timisoara Romania - CORRAlex MircescuNoch keine Bewertungen