Das könnte Ihnen auch gefallen
- The Toxicity of Caffein: An experimental study on different species of animalsVon EverandThe Toxicity of Caffein: An experimental study on different species of animalsNoch keine Bewertungen
- 12 Ivermectin Sensitivity in Collies Is Associated With A Deletion Mutation of The mdr1 GeneDokument7 Seiten12 Ivermectin Sensitivity in Collies Is Associated With A Deletion Mutation of The mdr1 GeneBerenice GranadosNoch keine Bewertungen
- tmp4F59 TMPDokument6 Seitentmp4F59 TMPFrontiersNoch keine Bewertungen
- IvermectinDokument5 SeitenIvermectinLFMSNoch keine Bewertungen
- Toxicidade Da Avermectina em Bovinos Com Menos de Trinta Dias de IdadeDokument10 SeitenToxicidade Da Avermectina em Bovinos Com Menos de Trinta Dias de IdadeErick AlvesNoch keine Bewertungen
- 2022 - Seguridad en Collies - Afoxolaner Solo en Combinación Con Milbemicina OximaDokument7 Seiten2022 - Seguridad en Collies - Afoxolaner Solo en Combinación Con Milbemicina OximaStephany GarciaNoch keine Bewertungen
- Molecular and Electrophysiological Characterization of GFP-Expressing CA1 Interneurons in GAD65-GFP MiceDokument11 SeitenMolecular and Electrophysiological Characterization of GFP-Expressing CA1 Interneurons in GAD65-GFP MiceFrontiersNoch keine Bewertungen
- Curine Inhibits Mast Cell Dependent Responses 2014 Journal of EthnopharmacoDokument7 SeitenCurine Inhibits Mast Cell Dependent Responses 2014 Journal of Ethnopharmacocds_tolucaNoch keine Bewertungen
- Url-2003 Parvovirus en Neuronas de GatoDokument5 SeitenUrl-2003 Parvovirus en Neuronas de GatocristianNoch keine Bewertungen
- Ijms 20 00107Dokument20 SeitenIjms 20 00107SRI RAHMAYUNINoch keine Bewertungen
- tmp90FE TMPDokument10 Seitentmp90FE TMPFrontiersNoch keine Bewertungen
- Journal of Neurochemistry - 2016 - Jalewa - Novel Incretin Analogues Improve Autophagy and Protect From MitochondrialDokument13 SeitenJournal of Neurochemistry - 2016 - Jalewa - Novel Incretin Analogues Improve Autophagy and Protect From Mitochondrialjalewa.jaishreeNoch keine Bewertungen
- NIH Public Access: p21 Protects "Super p53" Mice From The Radiation-Induced Gastrointestinal SyndromeDokument7 SeitenNIH Public Access: p21 Protects "Super p53" Mice From The Radiation-Induced Gastrointestinal Syndrome10sgNoch keine Bewertungen
- tmp5C3D TMPDokument10 Seitentmp5C3D TMPFrontiersNoch keine Bewertungen
- Pharmaceutics 08 00005Dokument13 SeitenPharmaceutics 08 00005abdalaahayoubNoch keine Bewertungen
- VEGFDokument12 SeitenVEGFlathifatulNoch keine Bewertungen
- Development Team: Zoology Molecular Genetics Transgenic Animals: ApplicationsDokument20 SeitenDevelopment Team: Zoology Molecular Genetics Transgenic Animals: ApplicationsBaikuntha SabarNoch keine Bewertungen
- The Central Sirtuin 1/p53 Pathway Is Essential For The Orexigenic Action of GhrelinDokument9 SeitenThe Central Sirtuin 1/p53 Pathway Is Essential For The Orexigenic Action of GhrelinJulia SCNoch keine Bewertungen
- Sem4a Insulin1995 JBCDokument4 SeitenSem4a Insulin1995 JBCAntonella Farabelli SuarezNoch keine Bewertungen
- Gano On BehaviourDokument4 SeitenGano On BehaviourBhavin VoraNoch keine Bewertungen
- Differential Transcripts TheDokument5 SeitenDifferential Transcripts TheGuhan KANoch keine Bewertungen
- Therapeutic Contraindications in Exotic PetsDokument14 SeitenTherapeutic Contraindications in Exotic Pets黃皓Noch keine Bewertungen
- Val (8) GLP-1 Rescues Synaptic Plasticity and Reduces Dense Core Plaques in APP/PS1 MiceDokument12 SeitenVal (8) GLP-1 Rescues Synaptic Plasticity and Reduces Dense Core Plaques in APP/PS1 Micegoonga_rishiNoch keine Bewertungen
- PDF Receptor Signaling in CaenorhabditisDokument9 SeitenPDF Receptor Signaling in CaenorhabditisDaniel Alejandro Herrera RamírezNoch keine Bewertungen
- J Jns 2004 12 011Dokument10 SeitenJ Jns 2004 12 011Viviana LunaNoch keine Bewertungen
- Fish & Shell Fish Immunology: Short CommunicationDokument8 SeitenFish & Shell Fish Immunology: Short CommunicationNur Maulida SafitriNoch keine Bewertungen
- Hepatorenal and Genotoxic Effects of Genetically Modified Quail Meat in A 90-Day Dietary Toxicity Study in MiceDokument7 SeitenHepatorenal and Genotoxic Effects of Genetically Modified Quail Meat in A 90-Day Dietary Toxicity Study in MiceDaniela CirnatuNoch keine Bewertungen
- The Effects of Sub-Chronic Exposure of Wistar Rats To The Herbicide Glyphosate-BiocarbDokument6 SeitenThe Effects of Sub-Chronic Exposure of Wistar Rats To The Herbicide Glyphosate-BiocarbFreddy A. ManayayNoch keine Bewertungen
- Clinical and Histological Evaluation of An Analogue of Palmitoylethanolamide, PLR 120 (Comicronized Palmidrol INN) in Cats With EosinDokument11 SeitenClinical and Histological Evaluation of An Analogue of Palmitoylethanolamide, PLR 120 (Comicronized Palmidrol INN) in Cats With EosinjenNoch keine Bewertungen
- Sem4a Insulin1995 JBC PDFDokument4 SeitenSem4a Insulin1995 JBC PDFAgus ParraNoch keine Bewertungen
- tmp490 TMPDokument11 Seitentmp490 TMPFrontiersNoch keine Bewertungen
- Ijpbs 2011Dokument8 SeitenIjpbs 2011Ramakrishna DevakiNoch keine Bewertungen
- Colarma,+1791 DefDokument8 SeitenColarma,+1791 Defhwyep160bNoch keine Bewertungen
- Apoptosis Inducing+Factor+DeficiencyDokument9 SeitenApoptosis Inducing+Factor+DeficiencyMariana Siqueira de OliveiraNoch keine Bewertungen
- Targeting The NLRP3 Inflammasome To Reduce Diet-Induced Metabolic Abnormalities in MiceDokument39 SeitenTargeting The NLRP3 Inflammasome To Reduce Diet-Induced Metabolic Abnormalities in Miceanirbanmanna88320Noch keine Bewertungen
- El Sayed2018Dokument11 SeitenEl Sayed2018Cindy Noor PradiniNoch keine Bewertungen
- Case Report Generalized Dermatophytosis in Six Yorkshire Terrier DogsDokument7 SeitenCase Report Generalized Dermatophytosis in Six Yorkshire Terrier DogsjenNoch keine Bewertungen
- Pemphigus: Nasal Dermatitis VulgarisDokument2 SeitenPemphigus: Nasal Dermatitis VulgarisMeuthia AlamsyahNoch keine Bewertungen
- The Rodenticidal Effect of Indomethacin: Pathogenesis and PathologyDokument12 SeitenThe Rodenticidal Effect of Indomethacin: Pathogenesis and PathologyNana Tabi GyekyeNoch keine Bewertungen
- IL31 Dermatitis AtipicaDokument8 SeitenIL31 Dermatitis AtipicaManu Ela GonzálezNoch keine Bewertungen
- PDFDokument12 SeitenPDFRidha Surya NugrahaNoch keine Bewertungen
- Protein Folding Proposal (Huntingtin)Dokument3 SeitenProtein Folding Proposal (Huntingtin)yuyupyupchemNoch keine Bewertungen
- Neuroprotective Dopamine - FullDokument7 SeitenNeuroprotective Dopamine - FullmorexyzNoch keine Bewertungen
- Doi10.1016-J.pnpbp.2008.03.024 2011Dokument8 SeitenDoi10.1016-J.pnpbp.2008.03.024 2011Mark BloomanNoch keine Bewertungen
- Flaxseed OilDokument36 SeitenFlaxseed OilKamal UddinNoch keine Bewertungen
- Fullerenes As An Effective Amyloid Fibrils Disaggregating Nanomaterialibqvb PDFDokument2 SeitenFullerenes As An Effective Amyloid Fibrils Disaggregating Nanomaterialibqvb PDFjeepdime3Noch keine Bewertungen
- Wolstenholme 2005Dokument11 SeitenWolstenholme 2005diijah678Noch keine Bewertungen
- Role of Ergot Alkaloids in The Immune SystemDokument17 SeitenRole of Ergot Alkaloids in The Immune SystemSyed Ali Akbar BokhariNoch keine Bewertungen
- 9243 FTPDokument7 Seiten9243 FTPnightrider92Noch keine Bewertungen
- Effect of Spinosad and Imidacloprid On Serum Biochemical Alterations in PDFDokument7 SeitenEffect of Spinosad and Imidacloprid On Serum Biochemical Alterations in PDFMekala LakshmanNoch keine Bewertungen
- Ravikanth3 PDFDokument7 SeitenRavikanth3 PDFMekala LakshmanNoch keine Bewertungen
- UprgenyDokument9 SeitenUprgenyJose Rafael Villafan BernalNoch keine Bewertungen
- Gene Expression Profiling in Granulosa Cells Under Levonorgestrel (2016)Dokument9 SeitenGene Expression Profiling in Granulosa Cells Under Levonorgestrel (2016)Iris MarRoNoch keine Bewertungen
- Aging and Disease: Connections To Sirtuins: ReviewDokument6 SeitenAging and Disease: Connections To Sirtuins: ReviewWwwanand111Noch keine Bewertungen
- 1 s2.0 S0898656822002972 MainDokument10 Seiten1 s2.0 S0898656822002972 Maindr. Nur'aini HasanNoch keine Bewertungen
- Of Galectin-3, f-Galactoside-Binding Lectin,: Expression and Function in Human Monocytes and MacrophagesDokument13 SeitenOf Galectin-3, f-Galactoside-Binding Lectin,: Expression and Function in Human Monocytes and MacrophagesMariana GarvilNoch keine Bewertungen
- tmp3D5 TMPDokument9 Seitentmp3D5 TMPFrontiersNoch keine Bewertungen
- Helicobacter Pylori: Upregulates Prion Protein Expression in Gastric Mucosa: A Possible Link To Prion DiseaseDokument6 SeitenHelicobacter Pylori: Upregulates Prion Protein Expression in Gastric Mucosa: A Possible Link To Prion DiseaseNadeem MalikNoch keine Bewertungen
- Glycobiology 1999 Dricu 571 9Dokument9 SeitenGlycobiology 1999 Dricu 571 9Romana MasnikosaNoch keine Bewertungen
- jfnr06 2 p047 054 Mlichova PDFDokument8 Seitenjfnr06 2 p047 054 Mlichova PDFTatum NilamsariNoch keine Bewertungen
- GMAT Reading Comprehension PracticeDokument84 SeitenGMAT Reading Comprehension PracticeDâu Tây100% (4)
- CellCommunication Assessment3Dokument6 SeitenCellCommunication Assessment3Michaela Maria GarciaNoch keine Bewertungen
- Ncbi Selulitis BulosaDokument3 SeitenNcbi Selulitis BulosaHarefa TirtaNoch keine Bewertungen
- DSE-03-U2 - Behavior & HealthDokument23 SeitenDSE-03-U2 - Behavior & HealthIndrashis MandalNoch keine Bewertungen
- 3 Cases About Personality DisorderDokument8 Seiten3 Cases About Personality DisorderJeane Rose MacauyagNoch keine Bewertungen
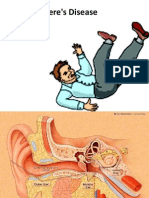
- Penyakit MeniereDokument25 SeitenPenyakit MeniereMuhammad Budiman Irpan BachtiarNoch keine Bewertungen
- HIp & Knee Extremity Notes Sorgenfrey Gindl 7.16.10Dokument116 SeitenHIp & Knee Extremity Notes Sorgenfrey Gindl 7.16.10joe joeNoch keine Bewertungen
- Challenges Faced by Families of Autistic Children-153Dokument5 SeitenChallenges Faced by Families of Autistic Children-153Zahoor Ahmad100% (1)
- Ahmed Proposal2Dokument34 SeitenAhmed Proposal2ahmedabdikerimNoch keine Bewertungen
- Mouse DissectionDokument6 SeitenMouse DissectionMauricio F. VillamarNoch keine Bewertungen
- Persuasive LetterDokument2 SeitenPersuasive Letterapi-341527188Noch keine Bewertungen
- Kinesiology Tape Guide - Physix Gear Sport PDFDokument85 SeitenKinesiology Tape Guide - Physix Gear Sport PDFTomas Delfiner100% (1)
- The Benefits of Cider VinegarDokument15 SeitenThe Benefits of Cider VinegarAldoSolsaNoch keine Bewertungen
- A Detailed Analysis of 16S Ribosomal RNA Gene SegmentsDokument10 SeitenA Detailed Analysis of 16S Ribosomal RNA Gene SegmentsCharith NikahetiyaNoch keine Bewertungen
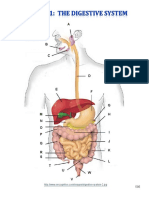
- Digestive System Charts 1Dokument14 SeitenDigestive System Charts 1api-306762527Noch keine Bewertungen
- PletizmografijaDokument29 SeitenPletizmografijaJelenaJankovNoch keine Bewertungen
- Trauma ToraksDokument30 SeitenTrauma ToraksMichael KwanNoch keine Bewertungen
- Waleed Hassan, 00551188, Vivid Institute of Occupational Safety & Health-940Dokument19 SeitenWaleed Hassan, 00551188, Vivid Institute of Occupational Safety & Health-940qadeesbhatti09511Noch keine Bewertungen
- TURP SyndromeDokument13 SeitenTURP SyndromeWindy Ari WijayaNoch keine Bewertungen
- Wound HealingDokument70 SeitenWound HealingSachin KpNoch keine Bewertungen
- Market Potential For Probiotic Nutritional Supplem PDFDokument7 SeitenMarket Potential For Probiotic Nutritional Supplem PDFshanmuga8801Noch keine Bewertungen
- The Sovereign Remedy: Touch-Pieces and The King's Evil. (Pt. I) / Noel WoolfDokument30 SeitenThe Sovereign Remedy: Touch-Pieces and The King's Evil. (Pt. I) / Noel WoolfDigital Library Numis (DLN)Noch keine Bewertungen
- Research Presentation Script I. Background of The Study & Introduction of Research TitleDokument3 SeitenResearch Presentation Script I. Background of The Study & Introduction of Research TitleSophia Adel RodriguezNoch keine Bewertungen
- BobackDokument4 SeitenBobackapi-324391171Noch keine Bewertungen
- Pharmacology of Drugs Acting On Renal SystemDokument20 SeitenPharmacology of Drugs Acting On Renal SystemBestha Chakri100% (2)
- Drug Use and MisuseDokument36 SeitenDrug Use and MisuseKristal Campbell100% (1)
- First Draft Code of Ethics For Midwives in EthiopiaDokument32 SeitenFirst Draft Code of Ethics For Midwives in Ethiopiamiadjafar463Noch keine Bewertungen
- Ferric Carboxymaltose (Hemojet) : Md. Arafat Hossain Brand Executive, SBMD, 01713354512Dokument5 SeitenFerric Carboxymaltose (Hemojet) : Md. Arafat Hossain Brand Executive, SBMD, 01713354512Heranmoy Kumar Bijon100% (1)
- Chapter One: Introduction To ArthropodsDokument11 SeitenChapter One: Introduction To ArthropodseyoelNoch keine Bewertungen