Das könnte Ihnen auch gefallen
- BjorklundDokument22 SeitenBjorklundapi-284232770100% (1)
- Cultural Evolution: How Darwinian Theory Can Explain Human Culture and Synthesize the Social SciencesVon EverandCultural Evolution: How Darwinian Theory Can Explain Human Culture and Synthesize the Social SciencesBewertung: 3 von 5 Sternen3/5 (1)
- Verbs, Bones, and Brains: Interdisciplinary Perspectives on Human NatureVon EverandVerbs, Bones, and Brains: Interdisciplinary Perspectives on Human NatureNoch keine Bewertungen
- Mesoudi CPhandbookDokument43 SeitenMesoudi CPhandbookAYDIN YAZGI (야즈그)Noch keine Bewertungen
- Cognitive Gadgets: The Cultural Evolution of ThinkingVon EverandCognitive Gadgets: The Cultural Evolution of ThinkingBewertung: 3.5 von 5 Sternen3.5/5 (3)
- Life Span Perspective PaperDokument3 SeitenLife Span Perspective PaperElbert WashingtonNoch keine Bewertungen
- Development Psychology (Lesson 1)Dokument4 SeitenDevelopment Psychology (Lesson 1)Maxine de la TorreNoch keine Bewertungen
- The New Biology: A Battle between Mechanism and OrganicismVon EverandThe New Biology: A Battle between Mechanism and OrganicismNoch keine Bewertungen
- Positive Youth Development1Dokument92 SeitenPositive Youth Development1MarcoNoch keine Bewertungen
- Evolutionary Psychology and Feminism - Final Published 2011Dokument20 SeitenEvolutionary Psychology and Feminism - Final Published 2011Rodrigo Caetano RahmeierNoch keine Bewertungen
- Notes For Unit One PSY105Dokument5 SeitenNotes For Unit One PSY105Thalia SandersNoch keine Bewertungen
- Culture and The Evolution Learning of Social: Mark V. FlinnDokument45 SeitenCulture and The Evolution Learning of Social: Mark V. FlinnfranciscoNoch keine Bewertungen
- Nature and NurtureDokument14 SeitenNature and NurtureCL PolancosNoch keine Bewertungen
- Homo BiologicusDokument3 SeitenHomo BiologicusCarolina Roldán0% (1)
- Biolog IDokument17 SeitenBiolog IWhina R AymaNoch keine Bewertungen
- Evolutionary - Psychology,.a.new - Paradigm JurnalDokument30 SeitenEvolutionary - Psychology,.a.new - Paradigm JurnalkarinatyasNoch keine Bewertungen
- E S 1989 Cosmides Tooby Psych CultureDokument21 SeitenE S 1989 Cosmides Tooby Psych CultureMatteo RiveraNoch keine Bewertungen
- Developmental Psychology - Simple PsychologyDokument6 SeitenDevelopmental Psychology - Simple PsychologyAngie MandeoyaNoch keine Bewertungen
- Cross-Cultural, Developmental Psychology: Integrating Approaches and Key InsightsDokument60 SeitenCross-Cultural, Developmental Psychology: Integrating Approaches and Key InsightsMonicaJelitaChiregarNoch keine Bewertungen
- Summary, Analysis & Review of Jonathan Haidt's The Righteous Mind by InstareadVon EverandSummary, Analysis & Review of Jonathan Haidt's The Righteous Mind by InstareadNoch keine Bewertungen
- 1996 Cross Cultural ApproachesDokument5 Seiten1996 Cross Cultural ApproachesBinat BatoolNoch keine Bewertungen
- Symbolic Analysis Cross-Culturally: The Rorschach TestVon EverandSymbolic Analysis Cross-Culturally: The Rorschach TestNoch keine Bewertungen
- The Body of Faith: A Biological History of Religion in AmericaVon EverandThe Body of Faith: A Biological History of Religion in AmericaNoch keine Bewertungen
- What Causes Human Behavior: Stars, Selves, Or Contingencies?Von EverandWhat Causes Human Behavior: Stars, Selves, Or Contingencies?Bewertung: 4 von 5 Sternen4/5 (1)
- Full Download Evolutionary Psychology The New Science of The Mind 4th Edition Buss Test BankDokument35 SeitenFull Download Evolutionary Psychology The New Science of The Mind 4th Edition Buss Test Bankalpen.logwoodp7aoer100% (29)
- Mogilski2016SocialDarwinism FinalDokument4 SeitenMogilski2016SocialDarwinism Finalმელაძე სოფოNoch keine Bewertungen
- What Is Developmental PsychologyDokument6 SeitenWhat Is Developmental PsychologyLehlohonolo MorokeNoch keine Bewertungen
- Multilevel Selection: Theoretical Foundations, Historical Examples, and Empirical EvidenceVon EverandMultilevel Selection: Theoretical Foundations, Historical Examples, and Empirical EvidenceNoch keine Bewertungen
- Developmental PsychologyDokument11 SeitenDevelopmental PsychologyGibreel SHNoch keine Bewertungen
- Evolutionary Psychology - Psychology - Oxford BibliographiesDokument20 SeitenEvolutionary Psychology - Psychology - Oxford BibliographiesErick Antonio Bravo ManuelNoch keine Bewertungen
- SociobiologyDokument9 SeitenSociobiologyपरी रायNoch keine Bewertungen
- Lesson 1 - Professional Development Lesson 1Dokument6 SeitenLesson 1 - Professional Development Lesson 1Cristel Anne A. LlamadorNoch keine Bewertungen
- Henric Hetal Five Mistake 11Dokument41 SeitenHenric Hetal Five Mistake 11Rok SilmariNoch keine Bewertungen
- Assessing Spirituality in a Diverse WorldVon EverandAssessing Spirituality in a Diverse WorldAmy L. AiNoch keine Bewertungen
- Clinical Evol PsychologyDokument25 SeitenClinical Evol PsychologyjnkjnNoch keine Bewertungen
- On Being Human: Distinctiveness, Dignity, Disability & DisposalVon EverandOn Being Human: Distinctiveness, Dignity, Disability & DisposalNoch keine Bewertungen
- Human Aggression-1997-Clin Psych RevDokument15 SeitenHuman Aggression-1997-Clin Psych RevKaran JagtapNoch keine Bewertungen
- Science in AnthDokument5 SeitenScience in AnthJm VictorianoNoch keine Bewertungen
- A Comparison of Biological and Cultural Evolution: Review ArticleDokument14 SeitenA Comparison of Biological and Cultural Evolution: Review ArticleSohail KirmaniNoch keine Bewertungen
- STS Module 1Dokument29 SeitenSTS Module 1Maria AngelicaNoch keine Bewertungen
- Eisler Levine 2002 Nurture Nature CaringDokument44 SeitenEisler Levine 2002 Nurture Nature CaringHomero SchlichtingNoch keine Bewertungen
- World's Largest Science, Technology & Medicine Open Access Book PublisherDokument18 SeitenWorld's Largest Science, Technology & Medicine Open Access Book PublisherLeona Fe MoldesNoch keine Bewertungen
- On Evolutionary EpistemologyDokument16 SeitenOn Evolutionary EpistemologysnorkelocerosNoch keine Bewertungen
- Attachment and Dynamic Practice: An Integrative Guide for Social Workers and Other CliniciansVon EverandAttachment and Dynamic Practice: An Integrative Guide for Social Workers and Other CliniciansBewertung: 5 von 5 Sternen5/5 (1)
- Ryff Et Al. (2021) - Eudaimonic and Hedonic Well-BeingDokument44 SeitenRyff Et Al. (2021) - Eudaimonic and Hedonic Well-BeingMarceloNoch keine Bewertungen
- Human Enhancements for Space Missions: Lunar, Martian, and Future Missions to the Outer PlanetsVon EverandHuman Enhancements for Space Missions: Lunar, Martian, and Future Missions to the Outer PlanetsKonrad SzocikNoch keine Bewertungen
- Beyond The Nature-Culture Dichotomy: A Proposal For Evolutionary AestheticsDokument11 SeitenBeyond The Nature-Culture Dichotomy: A Proposal For Evolutionary AestheticsCritica IunaNoch keine Bewertungen
- Evolutionary Psychology I (Guidebook)Dokument114 SeitenEvolutionary Psychology I (Guidebook)Viorel Bude93% (15)
- Political Descent: Malthus, Mutualism, and the Politics of Evolution in Victorian EnglandVon EverandPolitical Descent: Malthus, Mutualism, and the Politics of Evolution in Victorian EnglandNoch keine Bewertungen
- Reviewer in Dev PsychDokument5 SeitenReviewer in Dev PsychGrazielle Anne SancejaNoch keine Bewertungen
- HNC RC Alan Slater, Ian Hocking, and Jon Loose - Theories and Issues in ChildDokument30 SeitenHNC RC Alan Slater, Ian Hocking, and Jon Loose - Theories and Issues in ChildDea Novianda GeovanniNoch keine Bewertungen
- Psychiatry - Evolutionary Developmental Psychopathology - Ian PitchfordDokument280 SeitenPsychiatry - Evolutionary Developmental Psychopathology - Ian PitchfordAlberto Chicayban100% (1)
- Jung and Evolutionary PsychDokument20 SeitenJung and Evolutionary Psychbraeun6100% (1)
- From "Cultural Shock" To "ABC Framework": Development of Intercultural Contact TheoryDokument9 SeitenFrom "Cultural Shock" To "ABC Framework": Development of Intercultural Contact TheoryGeorgiana GattinaNoch keine Bewertungen
- The Evolution of Complexity by Means of Natural SelectionVon EverandThe Evolution of Complexity by Means of Natural SelectionBewertung: 3.5 von 5 Sternen3.5/5 (2)
- William Baum - Rules, Culture and FitnessDokument21 SeitenWilliam Baum - Rules, Culture and FitnessIrving Pérez MéndezNoch keine Bewertungen
- Dimensionalizing Cultures - The Hofstede Model in ContextDokument26 SeitenDimensionalizing Cultures - The Hofstede Model in ContextSofía MendozaNoch keine Bewertungen
- PersonalityDokument11 SeitenPersonalityNnoorNoch keine Bewertungen
- AdoptionDokument394 SeitenAdoptionanca mariaNoch keine Bewertungen
- Jones Universal Psychology Kinship LanguageDokument5 SeitenJones Universal Psychology Kinship LanguagegpdaveniaNoch keine Bewertungen
- Crocker SelffDokument18 SeitenCrocker SelffgpdaveniaNoch keine Bewertungen
- Giacinto ScelsiDokument3 SeitenGiacinto ScelsigpdaveniaNoch keine Bewertungen
- Welton Affectivity BodyDokument24 SeitenWelton Affectivity BodygpdaveniaNoch keine Bewertungen
- Hardoy Bs - As. 1955 1991Dokument26 SeitenHardoy Bs - As. 1955 1991gpdaveniaNoch keine Bewertungen
- Boyd Richerson Culture Part Human BiologyDokument24 SeitenBoyd Richerson Culture Part Human BiologygpdaveniaNoch keine Bewertungen
- Das Stigma Contagion DefectDokument18 SeitenDas Stigma Contagion DefectgpdaveniaNoch keine Bewertungen
- Lock Eclipse Gene Return DivinationDokument25 SeitenLock Eclipse Gene Return DivinationgpdaveniaNoch keine Bewertungen
- Richard Sociobiology Primate Field StudiesDokument26 SeitenRichard Sociobiology Primate Field StudiesgpdaveniaNoch keine Bewertungen
- Sapolsky Social Cultures Nonhuman PrimatesDokument16 SeitenSapolsky Social Cultures Nonhuman Primatesgpdavenia100% (1)
- Byrne Evolution Primate CognitionDokument28 SeitenByrne Evolution Primate CognitiongpdaveniaNoch keine Bewertungen
- Laland Animal Cultures DebateDokument6 SeitenLaland Animal Cultures DebategpdaveniaNoch keine Bewertungen
- Welton Affectivity BodyDokument24 SeitenWelton Affectivity BodygpdaveniaNoch keine Bewertungen
- A History of Membrane Transport and Bioenergetics PDFDokument383 SeitenA History of Membrane Transport and Bioenergetics PDFEdmilson RodriguesNoch keine Bewertungen
- Clinical Science MCQs - Sanfoundry PDFDokument2 SeitenClinical Science MCQs - Sanfoundry PDFSaqibKhanNoch keine Bewertungen
- Article: Contribution To The Knowledge of Ephemeroptera (Insecta) From Goiás State, BrazilDokument15 SeitenArticle: Contribution To The Knowledge of Ephemeroptera (Insecta) From Goiás State, BrazilErikcsen Augusto RaimundiNoch keine Bewertungen
- ProteinsDokument43 SeitenProteinsBilalNoch keine Bewertungen
- Toxicity of Clay Occulation of The Toxic Dino Agellate, Karenia Brevis, To Estuarine Invertebrates and SHDokument12 SeitenToxicity of Clay Occulation of The Toxic Dino Agellate, Karenia Brevis, To Estuarine Invertebrates and SHinousseNoch keine Bewertungen
- Producción y Caracterización de La Fenol Oxidasa de Scytalidium ThermophilumDokument7 SeitenProducción y Caracterización de La Fenol Oxidasa de Scytalidium ThermophilumDulzeKLFNoch keine Bewertungen
- Cysteamine - Full Profile - 170917Dokument11 SeitenCysteamine - Full Profile - 170917Walter MendozaNoch keine Bewertungen
- Corrections and Additions To Taxonomists' Glossary of Mosquito AnatomyDokument17 SeitenCorrections and Additions To Taxonomists' Glossary of Mosquito AnatomymokarahNoch keine Bewertungen
- Travel With An Ease While Not Putting Uneven Stress On Your Back - Caring Hands PhysiotherapyDokument1 SeiteTravel With An Ease While Not Putting Uneven Stress On Your Back - Caring Hands Physiotherapycaringhands physiotherapyNoch keine Bewertungen
- Esthetic Dentistry / Orthodontic Courses by Indian Dental AcademyDokument256 SeitenEsthetic Dentistry / Orthodontic Courses by Indian Dental Academyindian dental academyNoch keine Bewertungen
- 7 - IB Biology Required Practical 7 - Transpiration - IBDP - HL Lab Experiment GuideDokument16 Seiten7 - IB Biology Required Practical 7 - Transpiration - IBDP - HL Lab Experiment GuidemelindahofmannNoch keine Bewertungen
- MD II Material Pentru 24.03.2020Dokument5 SeitenMD II Material Pentru 24.03.2020Irina Panciu StefanNoch keine Bewertungen
- Anatomical Features of Bougainvillea (Nyctaginaceae) : Sarah ChewDokument7 SeitenAnatomical Features of Bougainvillea (Nyctaginaceae) : Sarah ChewPratistha ShauryaNoch keine Bewertungen
- Connect (4) New QuestionDokument18 SeitenConnect (4) New QuestionGeorge ElsabaNoch keine Bewertungen
- Test Bank For Introduction To Human Anatomy and Physiology 4th Edition by Pearl Solomon Isbn 10 0323239250 Isbn 13 9780323239257Dokument7 SeitenTest Bank For Introduction To Human Anatomy and Physiology 4th Edition by Pearl Solomon Isbn 10 0323239250 Isbn 13 9780323239257carwynquanh4tuozNoch keine Bewertungen
- TC18Dokument3 SeitenTC18Robert BarnesNoch keine Bewertungen
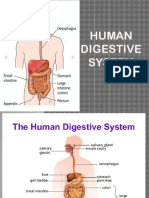
- Digestive-System-2 Grade 8Dokument39 SeitenDigestive-System-2 Grade 8Joshua Muego100% (2)
- Guia de Algas Bocas Del ToroDokument71 SeitenGuia de Algas Bocas Del ToroNohemi PaolaNoch keine Bewertungen
- MELD Related Files Liver MELD-PlusDokument45 SeitenMELD Related Files Liver MELD-PlusUKNoch keine Bewertungen
- 1 Determinant of MorbidityDokument55 Seiten1 Determinant of MorbidityGrace88 2016Noch keine Bewertungen
- Tu Eindhoven PHD ThesisDokument7 SeitenTu Eindhoven PHD ThesisHelpWritingAPaperManchester100% (2)
- 11 9700 23 2019 165347.inddDokument2 Seiten11 9700 23 2019 165347.inddAya TharwatNoch keine Bewertungen
- Vector NTI® Express SoftwareDokument184 SeitenVector NTI® Express SoftwareDrgemeNoch keine Bewertungen
- Nature Versus NurtureDokument58 SeitenNature Versus NurtureJade Harris' Smith100% (1)
- Pharmacology: A Career inDokument2 SeitenPharmacology: A Career insnikt7863443Noch keine Bewertungen
- The Beginnings of Modern Ornithology in Venezuela, Texera2002Dokument23 SeitenThe Beginnings of Modern Ornithology in Venezuela, Texera2002Jorge Luis Avila NúñezNoch keine Bewertungen
- Cerebrospinal FluidDokument3 SeitenCerebrospinal FluidlecturioNoch keine Bewertungen
- Understanding Immuno-Oncology - The Interplay Between TheDokument9 SeitenUnderstanding Immuno-Oncology - The Interplay Between Thenathalylara050590Noch keine Bewertungen
- Symbiotic Relationships Chart AnswersDokument2 SeitenSymbiotic Relationships Chart AnswersLiwayway Araza100% (1)