Das könnte Ihnen auch gefallen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- (Solutions Manual) Probability and Statistics For Engineers and ScientistsDokument51 Seiten(Solutions Manual) Probability and Statistics For Engineers and ScientistsAmy Parker100% (1)
- Physics of Cell Adhesion Failure and Human Diseases: SciencedirectDokument5 SeitenPhysics of Cell Adhesion Failure and Human Diseases: SciencedirectChukwuemeka JosephNoch keine Bewertungen
- The Effect of Hyperthermia On The PDFDokument18 SeitenThe Effect of Hyperthermia On The PDFChukwuemeka JosephNoch keine Bewertungen
- Mathematical Models Based On Transfer Functions To Estimate Tissue PDFDokument7 SeitenMathematical Models Based On Transfer Functions To Estimate Tissue PDFChukwuemeka JosephNoch keine Bewertungen
- Seminars in Cancer Biology: ReviewDokument32 SeitenSeminars in Cancer Biology: ReviewChukwuemeka JosephNoch keine Bewertungen
- Optical Properties of Fiber Optic Surgical Tips PDFDokument5 SeitenOptical Properties of Fiber Optic Surgical Tips PDFChukwuemeka JosephNoch keine Bewertungen
- A Theoretical Model Study of The Influence of Fluid Stresses On A CellDokument13 SeitenA Theoretical Model Study of The Influence of Fluid Stresses On A CellChukwuemeka JosephNoch keine Bewertungen
- Mutation Research/Reviews in Mutation Research: Franziska Van Zijl, Georg Krupitza, Wolfgang MikulitsDokument12 SeitenMutation Research/Reviews in Mutation Research: Franziska Van Zijl, Georg Krupitza, Wolfgang MikulitsChukwuemeka JosephNoch keine Bewertungen
- Seminars in Cancer Biology: ReviewDokument32 SeitenSeminars in Cancer Biology: ReviewChukwuemeka JosephNoch keine Bewertungen
- Akt Isoforms Regulate Intermediate Filament Protein Levels in Epithelial Carcinoma Cells - 1-S2.0-S0014579310000682-Main PDFDokument5 SeitenAkt Isoforms Regulate Intermediate Filament Protein Levels in Epithelial Carcinoma Cells - 1-S2.0-S0014579310000682-Main PDFChukwuemeka JosephNoch keine Bewertungen
- Laser Probe Ablation of Normal and Atherosclerotic PDFDokument11 SeitenLaser Probe Ablation of Normal and Atherosclerotic PDFChukwuemeka JosephNoch keine Bewertungen
- Turning The Heat On CancerDokument10 SeitenTurning The Heat On CancerChukwuemeka JosephNoch keine Bewertungen
- Optical Properties of Normal and Diseased Human Breast Tissues in The Visible and Near PDFDokument19 SeitenOptical Properties of Normal and Diseased Human Breast Tissues in The Visible and Near PDFChukwuemeka JosephNoch keine Bewertungen
- Er - YAG Laser Ablation of Tissue PDFDokument13 SeitenEr - YAG Laser Ablation of Tissue PDFChukwuemeka JosephNoch keine Bewertungen
- Characterization of Actuator Behaviors of Ferrogels Obtained From PDFDokument81 SeitenCharacterization of Actuator Behaviors of Ferrogels Obtained From PDFChukwuemeka JosephNoch keine Bewertungen
- Thermal Characteristics of Sapphire PDFDokument11 SeitenThermal Characteristics of Sapphire PDFChukwuemeka JosephNoch keine Bewertungen
- Malaysian Polymer Journal 6 1 39-50 PDFDokument12 SeitenMalaysian Polymer Journal 6 1 39-50 PDFChukwuemeka JosephNoch keine Bewertungen
- Physical Properties of Sapphire Fibretips For Laser PDFDokument6 SeitenPhysical Properties of Sapphire Fibretips For Laser PDFChukwuemeka JosephNoch keine Bewertungen
- PDMS Surface Modification Microvalves PDFDokument8 SeitenPDMS Surface Modification Microvalves PDFChukwuemeka JosephNoch keine Bewertungen
- Shape MemoryDokument16 SeitenShape MemoryFarooq MuhammadNoch keine Bewertungen
- An In-Vitro Study of The Effects of Temperature On Breast Cancer Cells Experiments PDFDokument8 SeitenAn In-Vitro Study of The Effects of Temperature On Breast Cancer Cells Experiments PDFChukwuemeka JosephNoch keine Bewertungen
- Cancer Facts & FiguresDokument1 SeiteCancer Facts & FiguresChukwuemeka JosephNoch keine Bewertungen
- Effect of Particle Size of Magnetite Nanoparticles On Heat Generating Ability Under Alternating Magnetic Field 2090 5025.110128Dokument4 SeitenEffect of Particle Size of Magnetite Nanoparticles On Heat Generating Ability Under Alternating Magnetic Field 2090 5025.110128Chukwuemeka JosephNoch keine Bewertungen
- Cancer and Radiation Therapy Current Advances and Future Directions PDFDokument7 SeitenCancer and Radiation Therapy Current Advances and Future Directions PDFChukwuemeka JosephNoch keine Bewertungen
- A Pharmaceutical Study of Doxorubicin-Loaded PEGylated Nanoparticles For PDFDokument10 SeitenA Pharmaceutical Study of Doxorubicin-Loaded PEGylated Nanoparticles For PDFChukwuemeka JosephNoch keine Bewertungen
- Ansys Linear Buckling PDFDokument5 SeitenAnsys Linear Buckling PDFJun Qian100% (1)
- Anti-Cancer Effect of Hyperthermia On Breast Cancer by Magnetite Nanoparticle-Loaded anti-HER2 Immunoliposomes PDFDokument1 SeiteAnti-Cancer Effect of Hyperthermia On Breast Cancer by Magnetite Nanoparticle-Loaded anti-HER2 Immunoliposomes PDFChukwuemeka JosephNoch keine Bewertungen
- Photo-Thermal Modeling of Plasmonic Heating of Concentric Nanoparticles PDFDokument6 SeitenPhoto-Thermal Modeling of Plasmonic Heating of Concentric Nanoparticles PDFChukwuemeka JosephNoch keine Bewertungen
- Combined Concurrent Photodynamic and Gold Nanoshell PDFDokument9 SeitenCombined Concurrent Photodynamic and Gold Nanoshell PDFChukwuemeka JosephNoch keine Bewertungen
- J-Integral ReportDokument26 SeitenJ-Integral ReportChukwuemeka JosephNoch keine Bewertungen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (119)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Design Considerations of High Voltage AnDokument114 SeitenDesign Considerations of High Voltage AnEL BRIGHLINoch keine Bewertungen
- Problems On Beams PDFDokument16 SeitenProblems On Beams PDFC j50% (6)
- The Mode of Eruptions and Their Tephra Deposits: Tetsuo K and Mitsuru ODokument8 SeitenThe Mode of Eruptions and Their Tephra Deposits: Tetsuo K and Mitsuru OAnggit Tri AtmajaNoch keine Bewertungen
- Optimum Penstocks For Low Head Microhydro Schemes - Alexander, Giddens - 2008Dokument13 SeitenOptimum Penstocks For Low Head Microhydro Schemes - Alexander, Giddens - 2008cbarajNoch keine Bewertungen
- Steel Castings Handbook: Supplement 2Dokument66 SeitenSteel Castings Handbook: Supplement 2Harshit AgarwalNoch keine Bewertungen
- TraysDokument23 SeitenTraysAmgadNoch keine Bewertungen
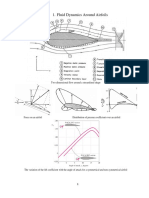
- Overview Aerodynamics 2017Dokument10 SeitenOverview Aerodynamics 2017marcoNoch keine Bewertungen
- Electronics Engg.: Detailed Solutions ofDokument52 SeitenElectronics Engg.: Detailed Solutions ofAshish ChoudharyNoch keine Bewertungen
- EPA 1668 A, Ag-2003Dokument129 SeitenEPA 1668 A, Ag-2003Karina Rondon RivadeneyraNoch keine Bewertungen
- April 2015-Vacuum Drying Chem Eng PDFDokument9 SeitenApril 2015-Vacuum Drying Chem Eng PDFAlvaro BarbozaNoch keine Bewertungen
- Analysis of Truss Using Abaqus SoftwareDokument7 SeitenAnalysis of Truss Using Abaqus SoftwareKarthick NNoch keine Bewertungen
- Mock Tests 20 20 Class 9 Science SampleDokument13 SeitenMock Tests 20 20 Class 9 Science SamplesrilathaNoch keine Bewertungen
- WRC 538 PDFDokument27 SeitenWRC 538 PDFsoojin gu25% (4)
- XI Maths - I Terminal ExamDokument2 SeitenXI Maths - I Terminal ExamPurisai Rajamani KumarNoch keine Bewertungen
- DWC ElecConduit Is 16205 P24 2018Dokument11 SeitenDWC ElecConduit Is 16205 P24 2018Vamsi Manoj60% (5)
- Sag Slurry PoolingDokument10 SeitenSag Slurry PoolingalgroneNoch keine Bewertungen
- Precision Thermometers CatalogueDokument44 SeitenPrecision Thermometers CataloguemarthaNoch keine Bewertungen
- Data Sheet Otdr Fho5000 enDokument2 SeitenData Sheet Otdr Fho5000 enJulio RoDriguezNoch keine Bewertungen
- Fundamentals of the Pickett Plot: Recognizing Reservoir PatternsDokument9 SeitenFundamentals of the Pickett Plot: Recognizing Reservoir PatternsAngelMeso100% (1)
- Weather in Kuttiadi - Google SearchDokument1 SeiteWeather in Kuttiadi - Google Searchsorry Its My StyleNoch keine Bewertungen
- How To Do Great WorkDokument64 SeitenHow To Do Great WorkMohan GuntiNoch keine Bewertungen
- Light SourcesDokument5 SeitenLight Sources123vidyaNoch keine Bewertungen
- P7 Revision QuestionsDokument4 SeitenP7 Revision Questionsapi-27344426Noch keine Bewertungen
- Welding Journal PDFDokument7 SeitenWelding Journal PDFraisalfiansyahNoch keine Bewertungen
- FTTH Accessories PDFDokument10 SeitenFTTH Accessories PDFdannyalcivarNoch keine Bewertungen
- Hawkins, Jaq - Elemental ChaosDokument3 SeitenHawkins, Jaq - Elemental Chaoscaligari2000Noch keine Bewertungen
- Determining Dielectric Constants Using A Parallel Plate CapacitorDokument5 SeitenDetermining Dielectric Constants Using A Parallel Plate CapacitorAhmAd GhAziNoch keine Bewertungen
- Java Meets Numerical AnalysisDokument2 SeitenJava Meets Numerical AnalysisbernasekNoch keine Bewertungen
- Lab 3 - Intro To DynamicDokument36 SeitenLab 3 - Intro To DynamicRacheal KirbyNoch keine Bewertungen
- Dosing Pump Innovata Drive ConceptDokument5 SeitenDosing Pump Innovata Drive ConceptgarpNoch keine Bewertungen