Beruflich Dokumente
Kultur Dokumente
IIntroducción
Hochgeladen von
Marlene GarcíaCopyright
Verfügbare Formate
Dieses Dokument teilen
Dokument teilen oder einbetten
Stufen Sie dieses Dokument als nützlich ein?
Sind diese Inhalte unangemessen?
Dieses Dokument meldenCopyright:
Verfügbare Formate
IIntroducción
Hochgeladen von
Marlene GarcíaCopyright:
Verfügbare Formate
I.
IIntroduccin II. Revisin de literatura 2.1. Aparato reproductor de la hembra 2.2. Fisiologa y endocrinologa de la hembra 2.2.1. Ovognesis. 2.2.2. Dinmica folicular. 2.2.3. Eje hipotlamo-hipfisis-gonadal. 2.2.4. Hormonas de la reproduccin. 2.2.5. Ciclo estral. 2.2.6. Anestro. 2.3. Fisiologa y endocrinologa del macho 2.3.1. Espermatognesis. 2.3.2. Control endocrinolgico. 2.3.3. Capacitacin espermtica. 2.3.4. Reaccin acrosomal. 2.3.5. Activacin e hiperactivacin. 2.3.6. Calidad del semen. 2.4. Inseminacin artificial. 2.5. Sin Est protocolos de sincro III. Resultados y discusin 3.1. Caso 1. Logan, E.F., Hdz, J.A., gutierrez, F. J. 2000. J. Anim. Sci. 22:123-130. 3.2. Ca 3.3. ca IV. Conclusin V. Lit cit
I Introduccin La reproduccin es el factor principal que limita la eficiencia de produccin del ganado vacuno ( ). Inseminacin artificial proporciona una tcnica econmicamente viable para introducir la gentica deseada en una manada, mientras que la sincronizacin del estro, ovulacin, o ambos proporciona una mano de obra ms eficiente manera de incorporar la AI en las prcticas de gestin. Sincronizacin de la AI tambin acorta la temporada de parto, lo que permite ms terneros que nazcan cerca del comienzo de la temporada de parto ( Larson et al., 2006 ). Tomando en cuenta lo anterior este trabajo pretendi abarcar los puntos clave de la inseminacin artificial en ganado de carne as como sus diferentes factores que lo alteran, tomando en cuenta desde la seleccin de los reproductores, mtodos ms viables de ordea del semen, como la evaluacin del semen morfologa como fsica (olor, color, consistencia etc.),y la conservacin del semen tambin la sincronizacin y deteccin del estro hasta la posterior inseminacin artificial del espcimen seleccionado (hembra). Estos puntos tienen la intencin de ayudar y mejorar en el aspecto reproductivo de la produccin agropecuaria en ganado de carne tomando en cuenta que en cada regin que intervienen distintos factores como son clima, flora, temperatura, el valor de la carga gentica del ganado. En cuestin, lo factores anteriores coadyuvara con esta prctica biotecnolgica como es el caso la inseminacin artificial tenga diferente valoracin en el aspecto de la eficiencia de esta prctica. No dejando afuera la buena realizacin de la practica desde la recoleccin del semen hasta la buena, correcta inseminacin artificial. La inseminacin artificial juega un papel muy importante en la reproduccin de grandes especies, demostrando ventajas que repercuten en la manera economa y el mejoramiento del hato, como por ejemplo: mejoramiento gentico, disminucin de costos, mayor sanidad animal. Introduccin de sangre a zonas inaccesibles y marginadas. Adems para tener un porcentaje alto de eficacia en la inseminacin artificial se tomaron en cuenta los siguientes puntos: seleccin de unos buenos reproductores, mtodos de las caractersticas de la recoleccin de semen, correcto manejo del semen en su congelacin y descongelamiento, hasta la correcta metodologa de la prctica de la inseminacin artificial. Para tener una mejor certeza en la inseminacin artificial se recomienda la sincronizacin de estros esto para tener un mejor control de los ciclos estrales de las vaca. Con la elaboracin de esta tesina se pretende llevar el conocimiento sobre las ventajas que da la inseminacin artificial en la produccin pecuaria dedicada a la produccin animal.
Realizar una recoleccin de datos sobre los diferentes mtodos que se conocen sobre la sincronizacin de estros y la inseminacin artificial en ganado de carne, teniendo especial inters
en las ventajas y desventajas de dichos mtodos, as tambin mostrando los pasos de las diferentes tcnicas y su correcta realizacin de la prctica de campo del mdico veterinario zootecnista.
II Revisin de literatura. 2.1. Aparato reproductor de la hembra. La vulva es el orificio externo del aparato reproductor, es la nica parte visible desde afuera de la vaca. Con 3 a 4 pulgadas de largo en el plano medio, la vulva est localizada inmediatamente debajo de la abertura externa del recto y de la cola. La vulva est compuesta de pliegues de piel gruesos y con comisuras agudas (epitelio estratificado escamoso, queratinizado) y cabellos que ofrecen una adecuada proteccin a las estructuras internas del rgano reproductor. La vulva al igual que otros rganos reproductivos durante el celo (tiempo de aceptacin del macho) es afectada por los efectos del estradiol incrementando la irrigacin (luce de color rojo); incrementando la humedad y el tejido se inflama (hinchada). Todas estas caractersticas son importantes cuando se evalan signos secundarios del celo. La vulva y el vestbulo son las nicas partes que comparten el tracto urinario y el tracto reproductivo. El cltoris posee un cuerpo que mide de 10 a 12 cm de largo y es sinuoso. El vestbulo es la primera estructura que se encuentra craneal a la vulva, mide 3-4 pulgadas de largo y1, 5 cm de ancho, cubierto por tejido escamoso, de epitelio no queratinizado. La abertura externa de la uretra est localizada en el piso del vestbulo. Caudal a esta estructura podemos encontrar un saco ciego (divertculo suburetral). La vagina es ancha, sus paredes son gruesas y su longitud es de 25 a 30 cm, durante la preez aumenta, se localiza craneal al vestbulo y se extiende cranealmente. Cubierta por epitelio estratificado escamoso no queratinizado. La vagina sirve de receptor del semen cuando se realiza la monta natural. El crvix con unas 4 a 5 pulgadas de largo y unas 2 pulgadas de ancho, es de suma importancia en la reproduccin bovina. En general el crvix es una rpida disminucin del tamao del tracto reproductor que sirve de proteccin del tero a la entrada externa de contaminantes que de otra manera fcilmente entraran desde la vagina. Durante la preez el crvix crea un tapn natural (tapn cervical) para crear un medio estril y seguro en el que vivir el feto. La ruptura de este tapn durante la preez que algunas veces puede suceder durante una inseminacin errnea, (de una vaca preada) puede provocar un aborto. La luz (lumen) del crvix es muy angosto y contiene una serie de pliegues de la mucosa que forma 3 o 4 anillos internos inclinados en direccin caudal. Hay algunas ramificaciones de estos principales o secundarios anillos que ayudan al transporte y reservorios del semen. Este gran nmero de anillos a su vez ayuda al crvix a expandirse durante el parto. Adems el crvix produce unas secreciones gruesas durante el metaestro, diestros y la preez y unas secreciones delgadas que son abundantes durante el celo que ayudan a facilitar el transporte y la lubricacin del semen.
El tero se localiza Craneal al crvix, se encuentra el tero, envuelto en el ligamento ancho del tero. El cuerpo mide de 3 a 4 cm de largo. Los cuernos miden de 35 a 40 cm de largo la parte libre
de los cuernos al inicio se dirigen ventral, craneal y lateral, para ser posteriormente caudal y dorsal, formando un espiral. Compuesto por epitelio ciliar columnar, el tero es el lugar donde se lleva a cabo la gestacin, es el responsable por brindar proteccin al feto y mantener una compleja comunicacin entre la madre y el feto. Las paredes del tero tienen numerosas funciones durante la gestacin. Tiene un tejido secretor que produce la leche uterina que sirve de nutriente para el embrin durante las primeras etapas de la gestacin. En el tero se pueden encontrar alrededor de 100 a 120 carnculas del tamao de un grano de maz distribuidas uniformemente en el endometrio. Estas carnculas sirven de punto de conexin para la placenta durante la preez. Las carnculas se unen ntimamente a los cotiledones para formar unas complejas y bien vascularizadas estructuras llamadas placentomas. Cada placentoma puede crecer hasta 2 pulgadas de ancho y sirve de unin de la placenta, de intercambio sanguneo que lleva nutrientes al feto y recoge los desechos del mismo para llevarlos a la orina de la madre. La placenta tambin se convierte en una poderosa glndula secretora que interacta con el endometrio y los ovarios para producir progesterona, estradiol, prolactina, y otras importantes hormonas. La pared uterina tiene una fuerte masa muscular que ayuda en la expulsin del feto al momento del parto y de las membranas fetales al poco tiempo despus del parto. Desde el punto de vista anatmico el tero puede dividirse en cuerpo del tero y dos cuernos. El cuerpo del tero es la primera parte del tero en direccin caudo-craneal y es la nica porcin compartida de las dos mitades derecha e izquierda del tero. Internamente tiene una medida de 2 pulgadas de largo antes de la iniciacin de la bifurcacin interna. Los oviductos son largos y menos flexuosas, miden de 20 a 25 cm cada oviducto (derecho e izquierdo) se convierte en la estructura que une los cuernos uterinos con el ovario, adems de ser el sitio donde se lleva a cabo la fertilizacin. El extremo craneal del oviducto presenta una abertura ancha y delgada en forma de embudo llamada fimbria la cual abraza el ovario y captura el vulo durante la ovulacin. Una vez que el vulo entra el oviducto, viaja y se deposita en la ampolla (la parte media del oviducto) esperando por el espermatozoide para llevar a cabo la fertilizacin. Si la fertilizacin ocurre el vulo fertilizado (embrin) viaja en direccin caudal a travs del istmo y la unin uterotubal para llegar al cuerno 3 a 4 das despus. Los ovarios son pequeos miden de 3 a 4 cm de longitud, 2,5 cm de ancho y 1,5 cm de grueso. El peso es de 15 a 20 gramos. Es de forma, agudo en su extremo uterino, no presenta fosa ovrica, contiene dos principales estructuras, el cuerpo lteo (produccin de progesterona) y los folculos que producen el estradiol y produccin de vulos en diferentes estados de madures. Aunque existen miles de vulos en los ovarios, todos ellos se formaron durante el desarrollo embrionario o fetal mucho antes del nacimiento. Durante el ciclo estral de una vaca un grupo de vulos compite por el desarrollo y la maduracin final (Folculos de Graff), ejerciendo dominancia durante cada onda folicular, pero bajo condiciones normales un solo folculo dominante dela ultima onda folicular puede ser ovulado.
2.2 Fisiologa y endocrinologa de la hembra.
2.2.1. Ovognesis. La hembra, desde que nace, posee miles de folculos primordiales en su ovario, muchos de los cuales estn destinados a permanecer como tales durante toda la vida o a sufrir atresia. Algunos, la minora, se desarrollarn hasta el estadio de folculo preovulatorio y liberarn el ovocito que contienen en el proceso de ovulacin. El folculo primordial est constituido por un ovocito primario procedente de las clulas germinales primitivas rodeado por una nica capa de clulas planas originadas a partir de los cordones celulares corticales. Estas clulas inhiben la maduracin del ovocito interrumpiendo la meiosis en la primera profase (estadio de diploteno difuso, caracterizado por la presencia de un ncleo prominente que recibe el nombre de vescula germinal y adems ejercen un efecto inhibidor sobre el crecimiento del ovocito.
En el folculo primario o preantral las clulas que rodean al ovocito se hacen cbicas y comienzan a dividirse, creando capas sucesivas en torno al mismo. Estas clulas estn comunicadas entre s por medio de uniones estrechas y constituyen el soporte nutritivo del ovocito. Sobre ellas aparece la lmina basal y entre el ovocito y las clulas foliculares se van depositando mucopolisacridos a modo de un anillo completo, constituyendo la zona pelcida. Por otra parte, las clulas fusiformes del estroma ovrico que se organizan alrededor de la membrana basal constituyen las clulas precursoras de la teca. Paralelamente, el efecto inhibidor que ejercan las clulas foliculares sobre el crecimiento del ovocito desaparece, aunque no ocurre lo mismo con la inhibicin de la meiosis. De este modo, el ovocito entra en una fase de intensa actividad sinttica y aumenta su volumen en ausencia de divisin celular, adquiriendo considerables reservas vitelinas. El paso de los folculos preantrales a folculos antrales inicia cuando el animal alcanza la pubertad. Las clulas de la granulosa, que han ido adquiriendo progresivamente receptores de membrana para FSH, comienzan a dividirse mitticamente al ser estimuladas por esta gonadotropina (liberada por el efecto GnRH). A su vez, estas clulas producen estrgenos que incrementan la actividad mittica de FSH; al aumentar el nmero y tamao de las clulas foliculares, aumenta el nmero de receptores de membrana para FSH y el proceso se auto estimula. La FSH, por otra parte, promueve tambin la produccin de lquido por las clulas foliculares lo que conlleva la aparicin del antro.
Al mismo tiempo, las clulas precursoras de la teca se diferencian completamente formando dos capas concntricas, la teca interna y la teca externa. La teca interna va adquiriendo
progresivamente una mayor vascularizacin y mayor riqueza en clulas epitelioides. La teca externa, formada por fibrocitos y clulas mioides con microfilamentos de actina y miosina, parece tener importancia en la contractilidad del folculo.
La liberacin de GnRH provoca a su vez una liberacin de LH por la adenohipfisis. En el folculo se establece un mecanismo conocido como "dos clulas, dos gonadotropinas" mediante el cual las clulas de la teca interna producen andrgenos que difunden a travs de la membrana basal hasta las clulas de la granulosa donde son transformados en estrgenos.
Durante las ltimas etapas de la maduracin folicular las clulas de la granulosa comienzan a expresar receptores para LH, que anteriormente solo se encontraban en las clulas de la teca. Este hecho permite al folculo responder al pico preovulatorio de LH que conducir a la maduracin del ovocito poco antes de la ovulacin. El pico preovulatorio de LH promueve tres cambios crticos en el folculo: la maduracin del ovocito, la ovulacin y la luteinizacin. Con respecto a la maduracin, el efecto del "inhibidor de la maduracin del ovocito" (OMI) intrafolicular es superado por la accin de la LH y el complejo cumulus-ovocito inicia una serie de cambios nucleares (alcanza la etapa de Metafase II) y citoplasmticos. Adems se produce un desacoplamiento progresivo entre las clulas del cumulusy entre stas y el ovocito, provocando la expansin o mucificacin del cumulus, propia de la maduracin del ovocito.
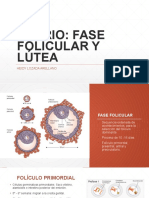
2.2.2. Dinmica folicular. Es la secuencia de crecimiento y atresia de folculos antrales que determinan la presencia de ondas de crecimiento folicular. 3 etapas: Reclutamiento: inicio de la onda. Un grupo de folculos antrales pequeos son reclutados por accin de las gonadotrofinas. La FSH estimula el crecimiento de estos que desarrollan receptores de LH en las clulas de la teca interna, lo que permite la sntesis de andrgenos, los cuales proveen el sustrato para que la FSH sinteticen estrgenos para la activacin de una aromatasa. Dura de 2 a 4 das. Seleccin: un folculo contina creciendo y se hace dominante sobre los dems, este es responsable de casi todo el estrgeno circulante en el plasma de la hembra. Por desarrollo de ms receptores o por secretar hormonas paracrinas inhiben el desarrollo de los dems.
Dominancia: el folculo seleccionado crece ms que los otros, que se atresian paulatinamente. En esta etapa hay 3 fases: 1. De crecimiento 2. Esttica: 1. Temprana. 2. Tarda. 3. Regresin: atresia en presencia de un cuerpo lteo funcional siendo alta la concentracin de P4 en la fase luteal. El estrgeno producido por el folculo dominante potencia la RAN de la P4 sobre GnRH, disminuyendo la frecuencia de pulsos de LH y FSH, atresiandoce el folculo. Posteriormente disminuye la concentracin de estrgeno, aumenta FSH y se inicia una nueva onda de crecimiento folicular. Foliculognesis. El folculo ovrico es la unidad estructural y funcional del ovario, proporcionando un ambiente adecuado para el crecimiento, eventual ovulacin del ovocito que contiene y formacin de un embrin a partir de la fecundacin (Cortvrindt & Smitz, 2001; Findlay et al., 2009). La foliculognesis es un proceso altamente selectivo donde usualmente solo un folculo asume dominancia y el destino del resto de los folculos es la atresia mediada por apoptosis; el mayor tipo de clulas que sufren este proceso son las clulas de la granulosa (Hsueh et al., 1994). Para Roberts (1971) y Knopf et al. (1989), la foliculognesis ocurre en estado fetal, animales prepberes y durante la gestacin. Un folculo primordial est compuesto por un ovocito con crecimiento detenido antes del nacimiento en la fase de diploteno de la profase I de la meiosis, rodeado por una sola capa de clulas de la pregranulosa (Nilsson et al, 2001). Segn Roberts (1971) y Fortune (2002), la formacin de folculos primordiales ocurre durante el periodo fetal. Asimismo, Roberts (1971) refiere que el crecimiento folicular, desarrollo a un folculo de Graaf y ovulacin ocurre solamente en hembras vacas despus de la pubertad y durante el ciclo reproductivo. Para Ginther (2000), el desarrollo y la atresia folicular estn reguladas por la sntesis de estradiol en las clulas de la granulosa, sin embargo para Faes et al. (2007), el estrgeno y la progesterona estn relacionadas con la calidad folicular. No obstante, para Basini et al. (1998), otros factores intraovricos no esteroidales tambin sintetizados por las clulas de la granulosa estn involucrados en la fisiologa ovrica. En las observaciones de Jolly et al. (1997), la LH estimula el AMPc en las clulas de la granulosa solamente en folculos con un dimetro mayor a nueve milmetros en taurinos y seis milmetros en cebunos, probando as, que los folculos mayores a este dimetro adquieren receptores para LH y, por tanto, capacidad ovulatoria. De esta forma, el tamao de los folculos influencia el potencial de desarrollo y el dimetro de los ovocitos (Witt & Kruip, 2001). Onda folicular. Ginther et al. (1989a), Adams et al. (1992), B et al. (1994) y Wiltbank et al. (1996), sugieren que el crecimiento de folculos bovinos ocurre en un patrn denominado ondas de crecimiento folicular.
Rajakoski (1960), basado en estudios histolgicos de ovarios, propuso la hiptesis de ocurrencia de dos ondas de crecimiento folicular durante el ciclo estral bovino, hiptesis reafirmada mediante ultrasonografa por Binelli (2000) y Borges et al. (2001), adems determinaron que durante cada onda de crecimiento folicular existe una poblacin de folculos pequeos, medianos y grandes en cada ovario, de los cules uno se torna dominante; segn Pierson & Ginther (1987), mediante un proceso de seleccin hasta el dimetro preovulatorio y generando la atresia de los folculos pequeos En la dinmica folicular ovrica pueden ocurrir de una a cuatro ondas de crecimiento folicular (Ginther et al., 1989a), es as que para Savio et al. (1988), en el 81% de los casos suceden dos ondas, tambin reportadas por Pierson & Ginther (1987), Binelli (2000) y Borges et al. (2001); sin embargo, Sirois & Fortune (1988), reportan que en el 80% de los casos ocurren tres ondas foliculares, halladas adems por Binelli (2000) y Borges et al. (2001). Existen diferencias en la dinmica folicular entre Bos taurus taurus (taurino) y Bos taurus indicus (cebuno), particularidad observada en el nmero de ondas de crecimiento folicular por ciclo estral, capacidad de secretar LH, rea del tejido luteal, dimetro folicular en el momento de la divergencia y en la ovulacin (Figueiredo et al., 1997; Pinheiro et al., 1998; Baruselli et al., 2007). En novillas Bos taurus indicus, Rhodes et al. (1995), Figueiredo et al. (1997), Viana et al. (2000) y Luiz (2002), reportan que la dinmica folicular es caracterizada por la presencia de dos ondas (33%) y tres ondas (57,1%), reportando hasta cuatro ondas por ciclo en Brahman, Nelor y Gyr. Savio et al. (1988), Sirois & Fortune (1988), Ginther et al. (1989a) y Wolfenson et al. (2004), refieren que en animales de la raza Holstein (Bos taurus taurus) predominan de dos a tres ondas por ciclo estral. Segn Carvalho et al. (2008), adems de la diferencia en el nmero de ondas foliculares, las hembras Bos taurus indicus reclutan mayor nmero de folculos por onda de crecimiento folicular que las hembras Bos taurus taurus: 33,43,2 versus 25,42,5, respectivamente. En hembras taurinas con dos ondas de crecimiento folicular el dimetro del folculo dominante es de 17,1 y 16,5 mm para la primera y segunda onda; en cebunas, los dimetros fueron de 11,3 y 12,1 mm, respectivamente (Ginther et al., 1989a; Figueiredo et al., 1997). Para B et al. (1994), entre el da uno a tres despus del estro emerge una onda de folculos que vara de 10 a 50 con un tamao de dos a tres milmetros, parte de los cuales continan creciendo hasta los cuatro y seis milmetros, de estos, entre dos y cinco siguen creciendo segn Ginther et al. (1996), a partir de la transicin de FSH a LH, donde ocurre la divergencia del folculo dominante (8,5 mm en taurinos y 6,2 mm en cebunos) (Baruselli et al., 2007), generando la regresin de los folculos menores, sin embargo, este folculo inicia su atresia luego de la fase esttica. Para Silcoux et al. (1993), en la primera onda de crecimiento folicular, la fase de crecimiento va desde la emergencia hasta cerca del octavo al dcimo da y la fase de regresin ocurre despus del dcimo da, para hembras que presentan dos ondas de crecimiento folicular, mientras que en las de tres ondas de crecimiento folicular se tiene del sexto al sptimo da de esttica y sptimo a octavo da de regresin.
Segn Savio et al. (1988) y Sirois & Fortune (1988), en animales que presentan ciclo estral de dos ondas de crecimiento folicular el reclutamiento de la primera onda es identificada en el da de la ovulacin, al da tres el folculo dominante est presente alcanzando el dimetro preovulatorio a los seis das, este folculo permanece esttico hasta el inicio de la segunda onda en el da diez despus de la ovulacin, donde se produce el folculo de Graaf y ovulatorio. Asimismo, refieren que en un ciclo de tres ondas foliculares la fase esttica del folculo dominante de la primera onda folicular es ms corta y la fase luteal es ms larga, en estos casos la tercera onda se inicia al da 16 y de esta se produce la ovulacin. Sin embargo, B et al. (1995), refieren que en el octavo da del ciclo estral (o sexto, por la variacin en el nmero de ondas) ocurre la emergencia de la segunda onda de crecimiento folicular y el proceso se reinicia; el folculo dominante de esa segunda onda regresiona (si hay tres ondas) o se torna folculo ovulatorio si sslo ocurren dos ondas. Variaciones en la dinmica folicular pueden deberse a factores como la dieta, manejo, produccin de leche, periodo de lactancia, y postparto (Ginther et al., 1996). La dieta puede afectar el patrn de ondas de crecimiento folicular, debido a que una nutricin pobre est asociada a bajas concentraciones de IGF-I circulante (Murphy et al., 1990), reduccin del dimetro del folculo dominante de todas las ondas y tambin reduce el tiempo de persistencia de este folculo durante la primera onda (Rhodes et al., 1995). Asimismo Rhodes et al. (1995), reportan que factores de tipo nutricional, de manejo y la poca del ao estn relacionados con la dinmica folicular que, para Badinga et al. (1994), involucran complejas interacciones neurohormonales del hipotlamohipfisis-ovarios. La pubertad. Es la etapa del desarrollo en la que el individuo adquiere la capacidad de liberar gametos viables y, por lo tanto, de reproducirse. La pubertad: Es un proceso dinmico Es gradual Es progresiva Incluye cambios somticos y celulares Precede a la madurez sexual Requiere, en general, el 60% del peso adulto del animal Durante el desarrollo perinatal, el sistema neurosecretor de la GnRH sufre una serie de cambios caractersticos del gnero, que dependen del ambiente esteroidal al que se vea sometido. Estos cambios generan patrones de secrecin de GnRH diferentes en machos y hembras.
Los machos solo liberan pulsos intermitentes y continuos de GnRH cuya secrecin se conoce como basal (centro tnico). Las hembras, adems de estos pulsos intermitentes pueden descargar grandes cantidades de GnRH en forma de pulsos peridicos lo que se conoce como pico preovulatorio de GnRH (centro cclico).
Los circuitos neuronales que regulan estos distintos tipos de secrecin en la hembra se conoces como centro tnico y centro cclico respectivamente. La retroalimentacin negativa que ejercen los estrgenos sobre la liberacin de GnRH acta en el centro tnico mientras que la positiva lo hace en el centro cclico. Las clulas nerviosas que componen al centro tnico reciben el nombre e integran la informacin proveniente de las pautas metablicas, medioambientales y endocrinas, determinando as una frecuencia y una amplitud de liberacin de GnRH. Los mecanismos neurobiolgicos que sincronizan la actividad de las neuronas de GnRH para generar una secrecin pulstil comprenden por un lado propiedades biofsicas propias de las clulas que les confieren un comportamiento rtmico intrnseco y por el otro la participacin de elementos externos aferentes como el neuropeptido y la norepinefrina. Teora del gonadostato: se comprob tempranamente que la capacidad de respuesta de la hipfisis y de las gnadas en los animales inmaduros era muy similar a la observada en los animales adultos, es decir cuando se le administraba GnRH a un individuo antes de la pubertad se logra la liberacin de LH por la hipfisis, igualmente cuando se aplicaban gonadotropinas exgenas, el ovario era capaz de desarrollar folculos en respuesta a la FSH y ovularlos por un estmulo de LH. Se sabe que los opioides suprimen la liberacin de GnRH durante el ciclo estral y la gestacin. El GABA inhibe la secrecin de GnRH en los animales prepuberes, al alcanzar la pubertad su concentracin se reduce lo que aparentemente se debe a la maduracin del transportador responsable del recambio de este neurotransmisor. Glutamato Estimula la secrecin de GnRH Su sensibilidad es ms alta al acercarse la pubertad Las neuronas secretoras de GnRH son inervadas directamente por neuronas glutaminergicas. Las neuronas que liberan glutamato en el hipotlamo se regulan parcialmente por estradiol. El cido glutmico (glutamato) es un aminocido, es un trasmisor.
La norepinefrina implicada en el control de la pulsatilidad de GnRH en roedores, el bloqueo de su liberacin retrasa la pubertad, su liberacin en el hipotlamo es sincrnica y pulstil con la GnRH, la secrecin de la norepinefrina se incrementa con la pubertad. La dopamina su tasa de recambio como su concentracin se incrementa con la pubertad en ratas. El neuropeptido parece estar relacionado con el mecanismo de la pubertad en pollos ratas y monos, su secrecin aumenta junto con la de GnRH en monos Rhesus hembra, en machos parece ser inhibitoria. La plasticidad del sistema nervioso y las relaciones que existen entre las neuronas, las clulas de la gla y las del endotelio vascular parecen jugar un papel importante en la presentacin de la pubertad. En el hipotlamo, los astrocitos secretan diferentes factores de crecimiento capaces de estimular la liberacin de GnRH indirectamente. El factor de crecimiento transformante (TGF) aumenta en la pubertad en roedores y primates. Estimula la secrecin de GnRH mediante la activacin de complejos de receptores que llevan a la produccin de prostaglandina E2 (PGE2), que se une a receptores especficos en las neuronas secretoras de GnRH. Los mecanismos que conducen a la pubertad y los eventos que la preceden pueden verse afectados por factores intrnsecos y extrnsecos al animal. Las concentraciones circulantes de elementos como el factor de crecimiento parecido a la insulina (IGF-I) o la leptina, contribuyen a la modulacin de los circuitos que desencadenan la liberacin de GnRH. Algunos centros nerviosos superiores como la glndula pineal o el bulbo olfatorio, monitorean los cambios medioambientales y los transmiten al sistema de control hipotalmico. 2.2.3 Eje hipotlamo-hipfisis-gonadal. El ciclo estral est regulado por una interaccin hormonal regida por el eje hipotlamo-hipfisisovario-tero. El hipotlamo forma la base del cerebro, y sus neuronas producen la hormona liberadora de gonadotrofina o GnRH. El GnRH, en la eminencia media, difunde a los capilares del sistema porta hipofisiario y de aqu a las clulas de la adenohipfisis en donde su funcin es estimular la sntesis y secrecin de las hormonas hipofisiarias, FSH y LH. La hipfisis est formada por una parte anterior o adenohipfisis y una posterior o neurohipfisis. La adenohipfisis produce varios tipos de hormonas, de las cuales la FSH y LH cumplen un papel relevante en el control neuroendcrino del ciclo estral. La FSH es la responsable del proceso de esteroideognesis ovrica, crecimiento y maduracin folicular, y la LH interviene en el proceso de
esteroideognesis ovrica, ovulacin, formacin y mantenimiento del cuerpo lteo. Estas hormonas son secretadas a la circulacin en forma de pulsos y son reguladas por dos sistemas, el tnico y el cclico. El sistema tnico produce el nivel basal circulante, siempre presente, de hormonas hipofisiarias las cuales promueven el desarrollo de los elementos germinales y endcrinos de las gnadas. El sistema cclico opera ms agudamente, siendo evidente por solo 12 a 24 horas en cada uno de los ciclos reproductivos de la hembra. El modo cclico tiene por funcin primaria causar la ovulacin. La neurohipfisis almacena la oxitocina producida en el hipotlamo. Esta hormona tiene varias funciones como son intervenir en el mecanismo del parto, bajada de la leche, transporte espermtico e intervendra en el proceso de lutelisis. Los ovarios son glndulas excrinas (liberan vulos) y endcrinas (secretan hormonas). Entre las hormonas que producen los ovarios podemos citar a los estrgenos, la progesterona y la inhibina. Los estrgenos, hormonas esteroideas, son producidos por el folculo ovrico y tienen acciones sobre los distintos rganos blanco como son las trompas de Falopio, el tero, la vagina, la vulva y el sistema nervioso central, en el cual estimulan la conducta de celo y el hipotlamo donde ejercen un "feed back" negativo sobre el centro tnico y positivo sobre el centro cclico. La progesterona, hormona esteroidea, es producida por el cuerpo lteo por accin de la LH. Los efectos de la progesterona se observan despus que el tejido blanco ha estado expuesto durante cierto tiempo a la estimulacin de los estrgenos. Esta preparacin por los estrgenos conduce a un efecto sinrgico. Esta hormona prepara el tero para el implante del embrin y para mantener la gestacin. A nivel hipotalmico ejerce un efecto feed back negativo sobre el centro tnico. La inhibina, hormona proteica, es producida por el folculo ovrico (clulas granulosas) e interviene en el mecanismo de regulacin de la secrecin de FSH. Ejerce un feed back negativo a nivel hipofisiario, produciendo una menor secrecin de FSH. TERO Produce la prostaglandina F2a (PGF2a), la cual interviene en la regulacin neuroendcrina del ciclo estral mediante su efecto luteoltico. Otras funciones son la de intervenir en los mecanismos de ovulacin y del parto. Hormonas de la reproduccin. Neurohormonas: actan directamente como hormonas en circulacin general. Neurosecreciones transportadas en pequeas vesculas envueltas en una membrana, hacia abajo va axones nerviosos hipotlamo-hipofisarios, mediante el flujo axoplasmico y se almacenan en la neurohipofisis hasta que se liberan a la circulacin. Oxitcina: tambin se produce en el cuerpo lteo. Tiene dos orgenes: hipotlamo y ovario. Relacionada con los patrones sexuales y con la conducta maternal acta tambin como neurotransmisor en el cerebro. En las hembras, la oxitcina se libera en grandes cantidades tras la distensin del crvix uterino y la vagina durante el parto, as como en respuesta a la estimulacin del pezn por la succin del beb, facilitando por tanto el parto y la lactancia. ACCION: 1) Sobre la musculatura lisa. 2) Contrctil, sobre clulas mioepiteliales de la glndula mamaria (eyeccin lctea).3) Para ayudar al parto o a la expulsin de placentas retenidas. 4)
Contrctil sobre las fibras longitudinales del miometrio permitiendo la expulsin del feto durante el parto y despus del parto la involucin uterina. 5) Liberada durante el coito (por macho y hembra) favoreciendo el transporte espermtico y el descenso del ovulo. 6) Induce la sntesis de PGF2 en la mucosa uterina participando en la luteolisis Hormonas reguladoras: actan en forma restringida sobre la hipfisis para regular su funcin. 6 estimuladoras, 3 inhibidoras. Hormona liberadora de gonadotrofinas GnRH: Es una hormona liberada por el hipotlamo cuyo centro de accin es la hipfisis. Es un decapptido que estimula la liberacin de gonadotrofina (hormona luteinizante o LH y folculo estimulante o FSH) por parte de la adenohipfisis. Por otro lado, la gonadotrofina poseen su centro de accin en las gnadas masculina y femenina. induce la sntesis y liberacin de FSH y LH por adenohipofisis. La secrecin est regulada por un oscilador neural, liberndose episdicamente a las venas portales hipofisarias imponindole un patrn de liberacin pulstil a la secrecin hipofisaria de gonadotrofinas que est ms marcado en LH que en FSH. Se utiliza para inducir y sincronizar ovulaciones, para tratar quistes ovricos. Hipofisarias: son de la adenohipofisis (la neurohipofisis no es una estructura glandular, sino solo un depsito de hormonas sintetizadas o producidas por el hipotlamo). Hormona folculo estimulante FSH: glicoprotena compuesta por dos subunidades, comn a la FSH, LH y TSH, y la especifica en su actividad biolgica. El periodo de vida media es de +/ - 2.5 horas. FSH regula el desarrollo, el crecimiento, la maduracin puberal, y los procesos reproductivos del cuerpo. FSH y LH actan de forma sinrgica en la reproduccin. ACCION: 1) Estimula peridicamente el desarrollo y crecimiento folicular, determinando las ondas de crecimiento folicular durante el ciclo estral (aumenta FSH= inicio de la onda, disminuye FSH= se selecciona el folculo dominante). Los folculos tienen receptores para FSH desde el estadio antral hasta el preovulatorio. 2) Junto con LH es responsable de la sntesis de estrgeno por los folculos en las clulas de la granulosa, una aromatasa que transforma sustancias androgenicas a 17 estradiol en la hembra tambin en las clulas de Sertoli (macho). 3) En macho acta en las clulas de sertoli, dentro de los tubos seminferos, donde estimula la sntesis de inhibina, estrgenos y protena transportadora de andrgenos. Es necesaria en la espermatogenesis. 4) Se utiliza en tratamientos de superovulacion para transferencias de embriones. Hormona luteinizante LH: glicoprotena, con caractersticas qumicas y tamao molecular muy similar a la FSH, producida por las clulas basofilas de la adenohipofisis, su vida media es de 30minutos. La liberacin de LH de la glndula hipfisis es regulada por la produccin pulstil de la hormona liberadora de gonadotrofinas (GnRH) proveniente del hipotlamo. Estos impulsos a su vez, estn sujetos a la retroalimentacin del estrgeno proveniente de las gnadas. ACCION: 1) Incrementa el flujo sanguneo en el ovario (efecto hiperhemico). 2) Estimula por si sola en la teca interna del folculo la sntesis de testosterona a partir de colesterol en la hembra; y en el macho acta sobre las clulas de Leydig. 3) Induce la formacin del cuerpo lteo y lo mantiene al
estimular la secrecin de progesterona por el cuerpo lteo. 4) Induce la ovulacin. 5) Se utiliza para sincronizar celos, para tratar quistes ovricos. Prolactina Pr: polipeptido secretado por las clulas acidofilas de la adenohipofisis. Estimula la produccin de leche en las glndulas mamarias y la sntesis de progesterona en el cuerpo lteo. ACCION: 1) Forma parte del complejo lactogenico hipofisario junto a ACTH y STH. 2) Induce el desarrollo del sistema de conductos y el crecimiento lbulo alveolar. 3) Favorece el pasaje de inmunoglobulinas hacia el calostro. 4) Inicia y mantiene la secrecin de leche. Hormona de crecimiento STH: protena secretada por las clulas acidofilas de la adenohipofisis. Acta a nivel del hueso, musculo, rin, tejido adiposo e hgado. La STH facilita el aumento de tamao de las clulas y estimula la mitosis, con lo que se desarrolla un nmero creciente de clulas y tiene lugar la diferenciacin de determinados tipos de clulas, como las clulas de crecimiento seo y los miocitos precoces. ACCION: 1) Estimula el crecimiento y el metabolismo. 2) Facilita el pasaje de aminocidos de la sangre a las clulas musculares.3) Es glucognica, aumentando los niveles circulantes de glucosa. 4) Estimula la actividad mittica en las epfisis de los huesos largos. 5) Aumenta la lipolisis y cidos grasos circulantes. 6) Incrementa la produccin de leche (galactopoyesis). 7) Estimula el crecimiento del tero, ovario y produccin de estrgenos. Tirotrofina TSH: glicoprotena secretada por las clulas basofilas de la adenohipofisis. ACCION: 1) La TSH hormona estimulante de la tiroides, aumenta la secrecin de tiroxina y triyodotironina por las glndulas tiroides produciendo la TSH en todas las actividades de las clulas glandulares tiroides. 2) Aumenta la protelisis de la tiroglobulina intrafolicular, con lo que aumenta la liberacin de hormona tiroidea hacia la sangre circulante y disminuye la substancia folicular misma. 3) Aumenta la actividad de la bomba de yodo que incrementa el ndice de captacin de yoduro en las clulas glandulares. 4) Aumenta la yodacin de la tiroxina y de su acoplamiento para formar hormonas tiroideas. 5) Aumenta el tamao y la funcin secretoria de clulas tiroideas. 6) Aumenta el nmero de clulas de las glndulas y hace que se transformen de cuboides en cilndricas. 7) La estimulacin elctrica del rea paraventricular del hipotlamo, aumenta la secrecin prehipofisiaria de TSH y en consecuencia aumenta la actividad de la glndula tiroides. Hormona estimulante de la corteza adrenal ACTH: protena producida por las clulas basofilas con un ritmo diurno bien establecido: el nivel plasmtico es mximo al amanecer y es mnimo al atardecer. estimula a las glndulas suprarrenales. ACCION: Regula la produccin de glucocorticoides por la corteza adrenal. Hormona melano estimulante: reguladora de los pigmentos cutneos. Es una protena, es una hormona vestigial en los mamferos.
NO hipofisarias: Gonadotrofina corionica equina CGe: glicoprotena con unidades y similares a FSH y LH, pero con mayor vida media. Secretada por las copas endometriales del tero equino, en las clulas trofoblasticas (entre el da 40 y 85 de la gestacin), se la encuentra en la sangre de donde se la extrae. ACCION: principalmente tipo FSH. Al estimularse el desarrollo de los folculos del ovario. Accin LH, ya que algunos folculos no ovulan y se luteinizan. Gonadotrofina corionica humana: sintetizada en el embrin y en la placenta de las mujeres embarazadas. La hCG durante el embarazo previene la desintegracin del cuerpo lteo en el ovario y por tanto mantiene la produccin de estrgenos, sumamente importantes para la preez, y adems interviene en la tolerancia inmunolgica durante la preez. Como la hCG se comienza a secretar despus de los primeros 6 das de preez, sta se puede usar como marcador de la gestacin. ACCION: tipo LH y algo FSH. Lactogeno placentario: es una hormona proteica similar a la prolactina, producida en el sincitiotrofoblasto la primera semana de la preez, alcanzando su concentracin mxima en el sexto mes (ltimo tercio de la gestacin). Mantiene el suministro constante de glucosa estimulando la lipolisis materna por manipulacin de las concentraciones y la sensibilidad materna a la insulina. Tambin aumenta el flujo de aminocidos hacia el feto. Hormonas gonadales. Hormonas gonadales esteroideas: secretadas por los ovarios, testculos, y corteza suprarrenal a partir del acetato. La placenta carece de enzimas para la sntesis de NOVO, pero puede sintetizar estrgeno y progesterona a partir del cortisol. Las hormonas esteroideas tienen una estructura qumica bsica comn: el ncleo est formado por el ciclo pentanoperhidrofenantreno, como el colesterol, difiriendo de las hormonas solo en las cadenas laterales unidas a este ncleo. Progestgenos: hormonas con un efecto similar a la progesterona, el nico progestgeno natural. Todos los dems progestgenos son sintticos. Progesterona: es secretada principalmente por el cuerpo lteo, y en menor medida por las clulas de la granulosa, en el folculo, poco antes de la ovulacin. Tambin por la corteza suprarrenal y por la placenta durante la preez. Es la hormona responsable del desarrollo de caracteres sexuales secundarios en una hembra, y sirve para mantener la preez. ACCION:1) Prepara al tero para la implantacin y mantenimiento de la preez: quietud uterina (inhibicin de la motilidad uterina) y aumento de glndulas secretoras del endometrio.2) Regula el ciclo estral: sinergiza con estrgeno induciendo estro y el comportamiento o receptividad sexual y elevadas concentraciones inhiben el estro o ciclo, y el pico ovulatorio de
LH. 3) Atresan folculos dominantes y NO inhibe el desarrollo de ondas de crecimiento folicular. 4) Estimula el desarrollo del tejido lbulo-alveolar, o secretor de la glndula mamaria durante la gestacin.5) Es anablica, contribuye a la ganancia de peso y deposicin de reservas en la madre a pesar del crecimiento fetal. 6) Cierra el canal cervical. 7) Se utiliza para sincronizar celos. Estrgenos: producidos por los ovarios y, en menores cantidades, por las glndulas adrenales. Inducen fenmenos de proliferacin celular sobre los rganos, principalmente endometrio, mama y el mismo ovario. Los estrgenos presentan su mayor concentracin los primeros 7 das de la menstruacin. La principal NATURAL es el 17-estradiol. Es sintetizada principalmente en las clulas de la granulosa de los folculos dominantes. Tambin en la placenta, corteza suprarrenal y clulas de Sertoli en machos. ACCION: 1) Caracteres sexuales secundarios de la hembra. 2) Junto con progesterona, induce a nivel del SNC, el comportamiento del celo y la libido. 3) Desarrollo del sistema de conductos mamarios. 4) Crecimiento endometrial, aumentando la irrigacin de los rganos de la reproduccin y provocando hipertrofia, aumentando el dimetro y la secrecin de la mucosa. 5) Durante el estro se observa, edema genital e hinchazn de vulva. 6) Estimula la sntesis de oxitocina en tero. 7) Regula la secrecin de LH y FSH. 8) Ovulacin. 8) Relajacin del crvix. 9) Estimula la asimilacin de calcio y la osificacin, impidiendo el desarrollo posterior. 10) Se utiliza para el control de los ciclos estrales, induccin de parto, expulsin de placentas, etc. Andrgenos: Los andrgenos son hormonas sexuales masculinas y corresponden a la testosterona, la androsterona y la androstendiona. Funcin principal es estimular el desarrollo de los caracteres sexuales masculinos. Son segregados por los testculos, en las clulas de Leydig, cuando son estimulados por LH. Tambin por los ovarios en la clulas de la teca del folculo, y por la corteza suprarrenal de las glndulas suprarrenales. ACCION: 1) Caracteres sexuales secundarios masculinos. 2) Comportamiento sexual masculino y la libido. 3) Crecimiento, desarrollo y actividad secretora de los rganos sexuales secundarios masculinos. 4) Funcionalidad de la tnica Dartos. 5) Estimula la ltima etapa de la espermatogenesis. 6) Regula RAN sobre hipotlamo e hipfisis a LH y FSH. 7) Se utiliza en animales como marcadores para la deteccin de celos. Hormonas gonadales NO esteroideas. Inhibinas: protena aislada de extractos testiculares y tambin en el lquido folicular. Su funcin principal es inhibir la secrecin de la gonadotrofina FSH. Tiene relacin con la testosterona. Producidas en las clulas de Sertoli en testculo y en folculos antrales (granulosa) en ovarios. Las inhibinas realizan RAN sobre hipotlamo e hipfisis, frenando la sntesis de FSH-Rh y FSH. ACCION: 1) Control en la secrecin de FSH. 2) Accin paracrina en el desarrollo folicular. Relaxina: polipeptido producido por el cuerpo lteo del ovario durante la preez. En algunas especies tambin por la placenta y el tero.
ACCION: 1) Dilatacin del crvix y vagina antes del parto. 2) Inhibicin de las contracciones miometriales. 3) Mejora la motilidad del esperma en el semen. Hormonas uterinas. Prostaglandinas: sustancias de carcter lipdico derivadas de los cidos grasos de 20 carbonos. Hormonas de accin paracrina en algunos casos endocrina no especfica, producidas por todos los tejidos del cuerpo. Tienen receptores de membrana que median respuestas rpidas y de corta duracin. ACCION: 1) Control de la presin sangunea, lipolisis, coagulacin de la sangre, secreciones gstricas, funcin renal y respiratoria. 2) Ovulacin. 3) Contracciones uterinas (Facilitan el parto, Facilitan el transporte de espermatozoides e Involucin uterina). 4) Luteotrofica. 5) Luteolisis. 6) Se utiliza para la sincronizacin de celos. Ciclo estral. En la vaca dura 20-21 das (+/- 18 das en vaquillonas). El periodo de CELO o receptividad sexual dura unas 18 horas, manifestndose en la vaca por actitudes como: Inquietud, mirada en accin de bsqueda. Orina con frecuencia. Pasividad ante la monta. Actitud homosexual (variable). Vulva tumefacta y hiperemia. Mucosidad que cae de la vulva.
La ovulacin ocurre de 10 a 12 horas despus de finalizado el celo y +/- 30 horas despus de iniciado. El da cero del ciclo es el da del CELO. Fases del ciclo estral: Fase folicular o de regresin luteal. PROESTRO: dura 3 o 4 das (corresponde al da 17-21 del ciclo), representa aproximadamente el 15%. Comienza con la lutolisis o regresin del cuerpo lteo (finaliza con el celo), cayendo abruptamente las concentraciones de P4 (progesterona), a las 24-36 horas. Esto sumado a los bajos niveles de E2, estimulndose la secrecin de LH que se libera en pulsos de mayor frecuencia y menor amplitud, en menor medida la secrecin de FSH, que estimula el desarrollo de un folculo
terciario o de Graaf, que secreta cantidades crecientes de E2 en forma de pulsos. Las clulas de la granulosa de los folculos antrales adquieren receptores para LH aumentando mucho la produccin de E2, producindose un pico de este 8 horas antes del inicio del celo.
Cuando se induce la lutolisis con PGF2, el folculo dominante solo puede ovular si se encuentra en las etapas de crecimiento o estadios tempranos (3-4 das); si el folculo se encuentra en etapas tardas o ha iniciado la atresia folicular, ovulara el folculo que se seleccione en la onda de crecimiento folicular siguiente (a los 4-6 das). La lutolisis se producir cuando el organismo no detecta la presencia de un embrin en el tero. Disminuyen los receptores endometriales para progesterona y aumentan para estrgeno, que inducen sntesis para receptores de oxitcica. La oxitcica de la neurohipofisis y el C.L aumenta el nmero de receptores de PGF2 y la sntesis en endometrio de PGF2, que por contracorriente llega al ovario y estimula la sntesis de oxitcica (RAP). Esta PGF2 disminuye el aporte sanguneo y la irrigacin. Provoca vasoconstriccin, hipoxia, disminuye el nmero de receptores para LH, aumenta la formacin de radicales superoxidos, disminuye la cantidad de progesterona y se produce LUTEOLISIS. El efecto luteolitico estara mediado por un aumento en la concentracin de calcio intracelular libre que induce apoptosis (al estimular endonucleasas que fragmentan el ADN, finaliza con el celo.
Periodo preovulatorio: ESTRO: el aumento en los niveles de E2, con muy bajos niveles de P4 circundante induce en el sistema nervioso modificaciones en la conducta animal caracterstica del periodo de receptividad sexual o celo. Los E2 ejercen RAP a nivel del centro cclico del hipotlamo, provocando la liberacin de una descarga muy grande de GnRh y LH (descarga preovulatoria de gonadotrofinas) aumentando la frecuencia y disminuyendo la amplitud de los pulsos, incrementndose los niveles basales de LH que alcanzan un pico mximo +/- a las 6 horas del inicio del celo (+/- 24 horas pre ovulacin). Este hecho desencadena la ovulacin, que ocurre a las 12 horas de terminado el celo (30 horas de iniciado). El pico de LH aumenta el riego sanguneo, disocia el cumulo ooforo, se sigue con la meiosis. La PGF2 estimula el plasminogeno que se transforma en plasma y junto a enzimas lisosomales liberadas por la PGF2, digieren la pared folicular, formndose el estigma. La PGF2 produce contracciones ovricas, produce la rotura del folculo y expulsin del oocito.
METAESTRO: el folculo se llena de sangre, transformndose en cuerpo hemorrgico que luego se luteiniza, por una serie de cambios quedando constituido por dos tipos de clulas latales las pequeas derivadas de la teca interna y las grandes derivada de la granulosa. Se va a empezar a formar el cuerpo lteo (las clulas foliculares se transforman en clulas latales) que va a secretar cantidades crecientes de progesterona (CL en formacin).
Fase luteal: DIESTRO: dura 10-12 das representa +/- el 60% del ciclo. Comprende desde que el cuerpo lteo es capaz de producir progesterona en respuesta de la LH la cual interviene en el desarrollo y mantenimiento del mismo. Este mecanismo se produce a travs de la formacin de AMP cclico y activacin de la protena quinasa A, con el consiguiente aumento en la produccin de progesterona. Si el oocito NO es fecundado el cuerpo lteo permanece funcional hasta el da 1619, luego del cual empieza a regresionar en preparacin de un nuevo ciclo estral.
Anestro.
Fisiologa y endocrinologa del macho La reproduccin en el macho es una funcin integrada, dependiente de la interaccin de seales hormonales y nerviosas entre el SNC, hipotlamo, hipfisis y testculo. Los testculos, al igual que los ovarios, producen y liberan clulas germinales y sintetizan hormonas esteroideas y peptdicas. Espermatognesis. Proceso que ocurre en los tubos seminferos y en el cual se originan los espermatozoides. Comienza en la pubertad y desde entonces se sucede de manera continuada. El proceso dura 60 das y tiene dos etapas: 1. Espermatocitogenesis: estimulada por FSH. Las espermatogonias (2n) se dividen mitticamente dando origen a 2 espermatocitos primarios (2n) cada una, que se dividirn meioticamente dando origen a 2 espermatocitos secundarios, estos posteriormente darn origen a 2 clulas llamadas espermatides (n). 2. Espermiogenesis: estimulada por testosterona las espermatides sufren una metamorfosis originando los espermatozoides.
Fase de diferenciacin o fase espermiognica: Consiste en la transformacin de las espermtidas en espermatozoides estructuralmente equipados para fertilizar al ovocito. Son necesarios niveles hormonales adecuados para el mantenimiento y la restauracin de la espermatognesis. Las hormonas necesarias para este proceso son: la testosterona, la FSH, la LH, la inhibina, los factores locales como el IGF-1, TGF-, oxitocina, vasopresina, opioides y citocinas. Control endocrinolgico. Por el eje hipotlamo-hipofisis-testculo. Factores ambientales sobre GnRH. GnRH producida por el hipotlamo en el rea hipofisiotropa, estimula la liberacin de FSH y LH por la adenohipofisis. La LH acta sobre las clulas intersticiales del testculo o de Leydig, posterior a una sensibilizacin sobre ellas de la FSH, e induce la sntesis y liberacin de testosterona. La FSH acta sobre las clulas de Sertoli en los tubos seminferos, transformando parte de la testosterona en estrgenos. El resto de testosterona va a los vasos sanguneos para dar los caracteres sexuales secundarios o a los tubos seminferos para funciones propias de la testosterona. Testosterona y estrgenos en la sangre actan sobre el hipotlamo e hipfisis frenando la produccin (RAN) de LH-RH (GnRH) y LH. Las inhibinas realizan RAN sobre el hipotlamo e hipfisis frenando la sntesis de FSH-RH (GnRH) y FSH. Capacitacin espermtica. La capacitacin normalmente ocurre en el aparato genital de la hembra, el sitio donde se inicia va a depender del sitio de depsito de los espermatozoides durante el coito. Los espermatozoides son capacitados gradualmente en poblaciones y no todos a la vez. La capacitacin espermtica consiste en el desarrollo funcional que sufre el espermatozoide, resultado de una serie de cambios o modificaciones estructurales y funcionales producto de su interaccin con las secreciones de la mucosa del aparato reproductor de la hembra. este proceso es necesario y provee al espermatozoide de las condiciones adecuadas para que se efecte la fertilizacin. En estas modificaciones se incluyen cambios bioqumicos y fisiolgicos en donde se alteran y eliminan sustancias que fueron integradas en la membrana plasmtica del espermatozoide durante su trnsito epididimal (por ejemplo, un factor descapacitante que tiene la propiedad de estabilizar y proteger al semen de sufrir la capacitacin y la reaccin acrosomal).
En preparacin para la reaccin acrosomal, durante la capacitacin, la membrana plasmtica del espermatozoide experimenta cambios en el contenido de fosfolpidos y colesterol. El contenido de
colesterol disminuye, lo que ocasiona que se incremente la permeabilidad y la fluidez de la membrana. Se ha propuesto que en la eliminacin gradual del colesterol participa la albmina presente en el fluido del aparato genital femenino. Adicionalmente, en la superficie del espermatozoide se presenta la eliminacin, modificacin y redistribucin de protenas y carbohidratos. Los cambios en los carbohidratos se ven reflejados en una disminucin de las cargas negativas de la superficie espermtica, debido a la modificacin de las sialoglicoprotenas, glicoprotenas y lpidos sulfatados. Lo que tambin ocasiona un aumento en la permeabilidad de la membrana y permite la fusin de membranas. En suma, las modificaciones en las propiedades fsicas de la membrana pueden influir en la movilidad, conformacin y activacin de las enzimas de la membrana y en el transporte de molculas. Esto ltimo por incrementar la permeabilidad de la membrana a los iones, entre ellos al calcio y/o pH. En particular, el calcio y el pH son involucrados en la capacitacin y en la reaccin acrosomal. El aumento del calcio y de protones tiene como resultado un decremento en la longevidad y una reaccin acrosomal prematura.
La remocin y/o la adsorcin de protenas a la superficie espermtica son generalmente reconocidas como un prerrequisito para la capacitacin. La capacitacin puede ser reversible, particularmente por la incubacin de los espermatozoides con las protenas del plasma seminal llamados factores descapacitantes. Por otra parte, el tiempo en el que se realiza la capacitacin es menor en los espermatozoides obtenidos de la cauda del epiddimo que en los eyaculados. Por otra parte, una hiperactivacin de la motilidad ha sido descrito en numerosas especies una vez que se realiza la capacitacin ya sea in vivo y/o in vitro; el espermatozoide aumenta la curvatura, la trayectoria y la amplitud flagelar, adems de un desplazamiento lateral de la cabeza. Diversos estudios han involucrado al AMPc y calcio en la regulacin. Se ha observado que una motilidad vigorosa requiere de AMPc, y que su accin es a travs de la fosforilacin de protenas asociadas con el axnema va protena cinasa A (PKA). La heparina es un componente utilizado en los medios para la capacitacin in vitro del espermatozoide del bovino. La heparina aumenta la sntesis de AMPc, eleva el pH y regula la capacitacin asociada con los cambios en la fosforilacin llevados por las protenas cinasas.
Reaccin acrosomal. El acrosoma es una estructura en forma de saco que ocupa la porcin anterior de la cabeza del espermatozoide recubriendo al ncleo y que se origina del Aparato de Golgi. El acrosoma est delimitado por una membrana continua, denominada como acrosomal externa (MAE) la adyacente a la membrana plasmtica y acrosomal interna (MAI), la cercana a la envoltura nuclear. La posterior del acrosoma recibe el nombre de segmento ecuatorial. El interior del acrosoma contiene una gran cantidad de enzimas hidrolticas, entre las que se encuentran: la acrosina, la hialuronidasa, la neuraminidasa, las fosfolipasas A, C y D, la catepsina D y la acetilglucosaminidasa, algunas de ellas necesarias para la degradacin de las envolturas del vulo.
La reaccin acrosomal (RA) es definida como una serie de eventos que involucran la liberacin del contenido acrosomal, el cual es un proceso de exocitosis regulada, ya que es inducida por dos molculas que han sido reconocidas como efectoras naturales de la RA; la progesterona que es secretada por las clulas de la granulosa y la glicoprotena ZP3 que es una molcula constitutiva de la zona pelcida (ZP). Sin embargo, la RA puede ocurrir de forma espontnea ya sea in vivo o in vitro. Morfolgicamente la RA verdadera ocurre en varios pasos: a) fusin de membrana plasmtica y la MAE en mltiples sitios con el inicio de la dispersin del contenido acrosomal, b) formacin y liberacin de las vesculas mixtas al medio extracelular junto con la liberacin completa de las enzimas y otros componentes contenidos en la matriz acrosomal; con la consecuente prdida del acrosoma, quedando as expuesta la MAI, c) fusin de la MP y un residuo de la MAI a nivel del segmento ecuatorial. El segmento ecuatorial del acrosoma no se ve involucrado en la vesiculacin de membranas durante la RA. Una vez que sucede la RA, los espermatozoides adquieren un nuevo dominio de fusin en la MP sobre la regin ecuatorial el cual puede ser reconocido por receptores presentes en la membrana plasmtica del vulo.
La capacitacin del espermatozoide es un requisito indispensable para que se de la RA. Si la capacitacin se da en ausencia de calcio la RA no se realiza de forma adecuada, pero si subsecuentemente se le adiciona, la RA se realiza de forma sincrnica. En muchas especies la ZP es capaz de inducir la RA. El papel de la ZP se ha estudiado de forma extensiva en los ratones, encontrndose a la protena ZP3 como responsable para de la adhesin e induccin de la reaccin acrosomal. La adhesin espermtica con la ZP involucra dos etapas: una adhesin primaria (una unin dbil) y una adhesin secundaria (una unin fuerte). La primera no es estrictamente especie-especfica y no requiere de capacitacin; en tanto que la secundaria es especie-especfica, requiere de la capacitacin espermtica y del calcio y no ocurre en los embriones o huevos activados. La cabeza de un espermatozoide capacitado expresa receptores especficos que son capaces de unirse a la ZP. La unin con la ZP estimula a los espermatozoides a sufrir la RA, en donde la MAE se fusiona con la MP dando como resultado la liberacin de las enzimas hidrolticas, como la acrosina, y la exposicin de un nuevo dominio de membrana, ambos resultados son esenciales para que el espermatozoide pueda fertilizar al vulo. La hialuronidasa liberada del espermatozoide con RA digiere la matriz de las clulas del cumulus y la acrosina ayuda al paso del espermatozoide a travs de la ZP (zona pelucida). Esta ltima produce una ranura con el ancho y la altura aproximada para que penetre la cabeza espermtica.
Activacin e hiperactivacin. Actividad.
El inicio y mantenimiento de la movilidad espermtica, estn asociados con la activacin de sistemas enzimticos, hay evidencias de que la actividad de la adenilato ciclasa aumenta durante la capacitacin, la cual eleva la concentracin intracelular de AMPc. Estos cambios favorecen la movilidad vigorosa y estimulan la actividad de las cinasas dependientes de este nucletido. La fosforilacin de protenas es inducida por AMPc e induce cambios siolgicos de la membrana, a travs de modicar las estructuras de las protenas presentes en ella. La actividad de la fosfolipasa A est asociada con los eventos de fusin caractersticos de la reaccin acrosomal. La accin de esta enzima sobre los fosfolpidos de la membrana provoca la acumulacin de lisofosfolpidos, que son compuestos capaces de producir fusin y lisis membranal (Flesch y Gadella, 2000). Hiperactivacin. Son los cambios en la movilidad espermtica, caracterizado en: a) aumento en el movimiento y exin del agelo, b) gran amplitud del desplazamiento lateral de la cabeza y c) trayectoria curva y tortuosa. Genera la fuerza requerida para la penetracin de la capa de clulas de la granulosa y la insercin inicial del espermatozoide con el ovocito (Tulsiani, 2006). Para inducirla se requiere Ca++ extracelular, elevacin intracelular de AMPc y disminucin del pH intracelular. La hiperactivacin del agelo se debe al Ca++ que se une a protenas jadoras en el brazo externo de la dinena (en la parte interna del agelo) induciendo el movimiento asimtrico (Inaba, 2003). Este Ca++ proviene de varias fuentes: a) Reservas intracelulares mediada por los receptores de inositol 1, 4,5 trifosfato (IP3), b) Mitocondria, c) o el que ingresa por sistemas de transporte Na+/H+ y Na+/Ca++ (Gadella et. al, 2008). Calidad del semen. CARACTERSTICAS MACROSCPICAS. VOLUMEN. Se mide directamente en la graduacin en mililitro (ml) del tubo de centrifuga o recipiente colector.
COLOR Y ASPECTO. Usualmente el semen del toro es de color blanco marfil y su aspecto vara entre acuoso, lechoso y cremoso, dependiendo de la cantidad de espermatozoides que contenga. El color amarillo limn que se observa en algunos toros es normal. Se debe a la presencia de riboflavina y es una
caracterstica hereditaria sin influencia alguna sobre la fertilidad. El color rojo o caf indica que contiene sangre fresca o sangre vieja.
CONCENTRACIN ESPERMTICA O DENSIDAD. La concentracin espermtica o densidad: es el nmero de espermatozoides por mm3 o cc. La concentracin normal del semen del bovino es de 800 a 1200 millones de espermatozoides por centmetro cbicos (ml), o sea, 0.8 a 1.2 millones por mm3. Tabla 1. Clasificacin densidad y color del semen. Clasificacin densidad y color del semen. Muy buena (MB): Semen cremoso, granular con 750 a 1,000 millones de espermatozoides/ml o ms. Buena (B): Semen lechoso con 400 a 750 millones de espermatozoides/ml. Suficiente (S): Semen semejante a leche descremada con 250 a 400 millones de espermatozoides/ml. Pobre (P): Semen translcido con menos de 250 millones de espermatozoides/ml.
CARACTERSTICAS MICROSCPICAS.
MOTILIDAD O MIVIMIENTO EN MASA. Se coloca una gota de semen de 5 mm de dimetro sobre un portaobjetos precalentado, y se observa el movimiento en masa de los espermatozoides usando microscopa de campo claro, el diafragma de campo cerrado y con magnificacin de 40x. Los factores que influyen en el movimiento en masa de los espermatozoides son la concentracin, el porcentaje de clulas con movimiento progresivo y la velocidad/vigor del movimiento de los espermatozoides. Si uno o ms de estos factores se encuentran comprometidos, el movimiento en masa se reducir. (Baracaldo Barth 2007).
Tabla 2. Clasificacin de motilidad en masa del semen. Clasificacin de motilidad en masa del semen. Muy bueno. Movimientos masivos muy marcados y rpidos. Bueno. Movimientos en masa aparente, pero moderados. Suficiente. Ondas en movimientos apenas apreciables. Pobre. No hay ondas, semen sin movimientos.
70-100% 50-69% 30-49% Menos de 30%
Tabla 3. Clasificacin descriptiva del movimiento en masa. Clasificacin descriptiva del movimiento en masa. Muy bueno (MB) Remolinos oscuros y rpidos Bueno (B) Remolinos lentos Regular (R) Sin remolinos, pero movimientos visible de clulas individuales Pobre (P) Poco o nada de movimientos celular individual
MOTILIDAD INDIVIDUAL PROGRESIVA. Se coloca un volumen de 5 a 7 l de semen sobre un porta objetos nuevo y precalentado creando una gota de aproximadamente 3 a 5 mm de dimetro, que luego se cubre con una laminilla cubreobjetos.
La muestra es observada con microscopia de contraste de fase y con aumento de 200-400x, determinado el porcentaje de clulas espermticas con movimientos progresivo lineal. Si el semen est muy concentrado, la muestra puede diluirse con una solucin amortiguadora o diluyente de semen antes de colocar el cubreobjetos.
Tabla 4. Evaluacin descriptiva de la motilidad individual. Evaluacin descriptiva de la motilidad individual. Muy buena (MB) 80-100% con motilidad progresiva Buena (B) 60-79% con motilidad progresiva Regular (R) 40-59% con motilidad progresiva Pobre (P) 40% con motilidad progresiva
pH. Se mide con el papel tornasol o con el potencimetro. El pH normal de semen de bovino es de 6.9 (6.4-7.8).
MORFOLOGA. Para observar la cantidad de espermatozoides vivos y las anormalidades, generalmente se mezcla el semen con colorantes y se realiza un frotis. En otras tcnicas, primero se hace un frotis del semen y luego se tie la laminilla. Existen gran cantidad de tinciones, como: eosina-nigrosina, rojo de Bengala-Azul Victoria, Wells, Williams, Giemsa, tinta china, Fenicada, violeta de metilo, anilina y coloracin de Karras. Tambin es posible usar preparaciones lquidas (solucin de Hancok)
mezcladas con el semen y observarlas al microscopio de contraste de fase. Por lo comn, se cuenta 300 espermatozoides en varios campos y se anotan los alterados y el tipo de alteracin. Inseminacin artificial. En esta seccin se describe la tcnica de inseminacin ms usada, la de fijacin cervical transrectal, por ser la ms eficiente. Otros mtodos que se utilizaron en algn momento son el vaginal y el del vaginoscopio. Se realizaron comparaciones entre los tres mtodos, tras las cuales el mtodo recto-vaginal o de fijacin cervical transrectal ha demostrado ampliamente ser el ms eficiente, por su elevado ndice de no retorno al celo, adems de ventajas sanitarias y prcticas. DESCRIPCIN DEL MTODO DE FIJACIN CERVICAL TRANSRECTAL.
El objetivo de estas lneas es interiorizar al lector acerca de los detalles ms importantes a tener en cuenta en el momento de la ejecucin de la tcnica ms adecuada para inseminar vacas. Todo el equipo utilizado para la inseminacin debe estar perfectamente limpio, los tubos de dilucin y pipetas o vainas deben estar esterilizadas o utilizarse elementos descartables, ya que son econmicos y desechables. Se utilizan igualmente guantes descartables, aunque algunos tcnicos prefieren usar de ltex El tcnico en el campo debe mantener sus instrumentos bien limpios y desinfectados, el delantal limpio y en buen estado. Cuando se usa semen congelado y tubos, pipetas o vainas descartables, no es preciso limpiar el equipo inseminador. Todo inseminador debe tener un tacho de desperdicios para el equipo usado; no hay nada peor que los catteres, guantes y material desechables en general, desparramados en el rea de trabajo. Ya se mencion que el mtodo ms eficiente es el recto-vaginal; recurrir al vaginoscopio o depositar el semen en vagina, constituye una impericia de principiante. La tcnica de fijacin cervical, tal como se muestra en la Figura N 15, es de empleo universal y la aplicacin es la misma ya sea que se use semen fresco o congelado, almacenado en pajuelas o pastillas.
Imagen 1. Proceso de inseminacin. Procedimiento: Los pasos a seguir para inseminar a la vaca segn la tcnica mencionada son los siguientes: Identificar correctamente al animal y sujetar al mismo. Limpiar la vulva y partes adyacentes con agua y posteriormente secar con toallas de papel. Preparar el semen y acondicionarlo en el instrumental de inseminacin. Ubicar el pistolete de inseminacin en el vestbulo de la vagina, separando previamente los labios vulvares y orientando el instrumento hacia el techo de la vagina. Colocar la mano enguantada (izquierda) con los dedos en forma de cua e introducirla suavemente en el recto, venciendo la resistencia del esfnter anal, eliminando el estircol de la ampolla rectal. Insertar el catter o pistolete de inseminacin con la mano libre (derecha) hacia el fondo de la vagina y guiarlo hasta el cuello uterino. Sujetar firmemente ste y enfrentar la punta del instrumento con la entrada del cuello. Manipular el instrumental de manera que pase a travs de los pliegues cervicales hasta la porcin posterior del canal. El pistolete o catter no debe penetrar profundamente en el tero, ya que algunas veces puede producir infeccin o bien el semen ser depositado en el cuerno contra-lateral al ovario ovulante. Generalmente los principiantes tienen dificultades al introducir por primera vez la pipeta en el
cuello uterino. Se necesita prctica y paciencia. En algunas vacas es imposible pasar completamente el instrumento a lo largo de todo el crvix y simplemente se deposita tan profundamente como se avanz en el mismo. La capacitacin del tcnico es un aspecto fundamental (5). La deposicin del semen se realiza en el cuerpo del tero. Una vez encontrado el sitio de deposicin, expeler el semen lentamente y retirar el instrumento.
Imagen 2. El lugar correcto para la deposicin del semen es el cuerpo uterino. Descartar el equipo usado, lavarse las manos y limpiar y desinfectar las botas y guantes si son de ltex. Completar los formularios y registros de reproduccin. Protocolos de sincronizacin. Segn Patterson et al (2000) la evolucin de los mtodos para el control del ciclo estral en la vaca, puede ser ordenado en 5 fases distintas. La primera comprende todas investigaciones con el sentido de prolongar la fase ltea a travs de la administracin de progesterona exgena. Con el tiempo estos mtodos pasaron a contar con una asociacin de estrgenos y gonadotropinas. La tercer fase esta caracterizada por la utilizacin de prostaglandinas con el fin de acortar la fase ltea, la cuarta fase seria aquella en la que fueron desarrollados los mtodos con la asociacin de progestgenos y prostaglandinas. La denominada quinta fase surgi por estudios ms recientes de las ondas foliculares que mostraron que el control del ciclo estral en la vaca requiere la manipulacin no solo de la fase ltea sino tambin del crecimiento folicular. Dentro de las ventajas de la sincronizacin de estros en bovinos podemos citar las siguientes: Concentracin de animales en estro en un corto periodo
Racionalizacin de la IA principalmente en vacas de carne. Concentracin y reduccin del periodo de paricin. Manejo de los alimentos disponibles de acuerdo con la poca del ao y las categoras de animales. Facilitar la formacin de test de evaluacin zootcnica para posibilitar la compra de individuos con intervalos reducidos entre los nacimientos. Registro de los terneros, facilitando las prcticas de manejo y comercializacin. Los principales factores limitantes a una mejor expansin en la utilizacin de los protocolos de sincronizacin de los protocolos de sincronizacin de celos y ovulacin en vacas, est asociado relativamente a los altos costos de las hormonas; desconocimiento por parte de los tcnicos sobre los mecanismos fisiolgicos que rigen la funcin reproductiva de la vaca, situaciones frecuentes en nuestro sistema de produccin con periodos de restriccin alimentara, as como una pequea reduccin de la fertilidad de los animales despus de los celos inducidos. Cuando se va a implementar un programa de sincronizacin tenemos que caracterizar al grupo de animales que sern tratados. Esta clasificacin se da bsicamente considerando si se trata de vaquillonas o vacas con cra al pie y el estado del ovario. Determinados protocolos que pueden ser utilizados en vacas o vaquillonas cclicas, son inadecuados en hembras acclicas. Actualmente existen 2 grupos de preparaciones hormonales disponibles en el mercado que pueden ser utilizadas para sincronizar celos en los bovinos: 1-Progestgenos que tienen como efecto principal un bloqueo hipotlamo-hipofisiario simulando una fase ltea. 2-Prostaglandinas y sus anlogos que actan como agente luteoltico sobre el cuerpo lteo. 2.5.1 Protocolos con progestgenos.
Bloqueo a travs de la administracin de MGA (Acetato de Melengestrol) Existen variaciones en cuanto a los protocolos que utiliza el MGA. En 1994 Anderson y Day propusieron una administracin diaria de MGA durante 14 das. Luego se verifico que reduciendo el periodo de tratamiento se obtena mayor fertilidad. Actualmente los protocolos ms recomendados, preveen la administracin de 0,5mg de MGA por cabeza por da durante 7 das mesturado con una racin. En el sptimo da luego de la suspensin del MGA se administra prostaglandina (dosis recomendada por el fabricante) provocando la lisis del cuerpo lteo de animales que ya estaban ciclando al comienzo del tratamiento. Cuatro das despus de la aplicacin de prostaglandina, con el objetivo de inducir la ovulacin o luteinizacin
folicular, se administra GnRH. La inseminacin artificial es realizada luego de la deteccin de celo, 48 a 96 hs posteriores a la aplicacin de prostaglandina. Este protocolo est indicado principalmente para vaquillonas prximas al inicio de la pubertad o ya pberes y en vacas acclicas posparto. Bloqueo a travs del implante subcutneo de Norgestomet El Norgetomet es un potente progestgeno sinttico que es utilizado de forma de implante subcutneo el cual contiene impregnado 3 mg (Crestar) del principio activo. El primer implante que surgi en el mercado fue el Syncromate B, el cual contiene 6 mg de Norgestomet. Estos implantes se aplican en la cara dorsal de la oreja del animal, permaneciendo por 9 das. Cuando se coloca el implante se administran 5mg de Valerato de Estradiol y 3 mg de Norgestomet, el primero para promover la luteolisis de un eventual cuerpo luteo y sincronizar la onda de crecimiento folicular, y el segundo con el intento de promover altas concentraciones de Norgestomet en el inicio del tratamiento, promoviendo con esto de inmediato el bloqueo hipotalmico-hipofisiario. En caso de posibles animales cclicos del grupo tratado, se recomienda cuando se retira el implante la aplicacin de una dosis de prostaglandina. Para vacas, las cuales se sabe que estn acclicas, se indica en este momento la administracin de 400 a 700 UI de eCG. La inseminacin artificial se realiza en un tiempo predeterminado, aproximadamente 50 hs posteriores al retiro del implante. Bloqueo a travs de la utilizacin de dispositivos intravaginales Actualmente en el mercado se encuentran disponibles diferentes tipos de dispositivos intravaginales los cuales contienen concentraciones variadas de progesterona, como por ejemplo tenemos: CIDR-B (1,9 g de progesterona), PRID (1,55 g de progesterona), DIB (1 g de progesterona), DISPOCEL (1 g de progesterona), etc. Uno de los ms utilizados es el CIDR-B. Este dispositivo consta con un implante en forma de T de silicona con un molde de nylon impregnado con 1,9 g de progesterona. La mucosa vaginal absorbe aproximadamente 0,5 a 0,6 mg de progesterona al da, determinndose esta forma el bloqueo hipotalmico-hipofisiario. El dispositivo es introducido en la cavidad vaginal a travs de un aplicador semejante a un especulo que mantiene las extremidades de la T aproximadas a manera de facilitar su introduccin. La extremidad distal del CIDR contiene un filamento de nylon que al final del periodo de utilizacin sirve para la remocin del dispositivo por traccin.
El protocolo tradicional de utilizacin del CIDR preconiza la permanencia del dispositivo en la cavidad vaginal por un periodo de 9 das. En el da de aplicacin del dispositivo se recomienda la aplicacin intramuscular de 2 mg de Benzoato de Estradiol, principalmente con el objetivo de sincronizar el crecimiento folicular. En este mismo momento se administran 50 mg de progesterona va intramuscular para auxiliar el inicio del bloqueo. Para grupo de animales cclicos que sern tratados, se hace necesaria la aplicacin de prostaglandina al momento de la retirada de los dispositivos. Como auxiliar del desencadenamiento de la ovulacin, es de utilidad la administracin de 1 mg de Benzoato de Estradiol intramuscular en el dcimo da del protocolo, realizando la inseminacin artificial a tiempo fijo cercano a las 50 horas posteriores a la retirada del dispositivo. Existen protocolos que preveen la sustitucin de Benzoato de Estradiol por dos aplicaciones de 100 mcg de GnRH, siendo la segunda realizada en el momento de la inseminacin artificial. En vacas que estn amamantando terneros con gran probabilidad de que se encuentren en estado de acclia, al momento de retirar el CIDR, en vez de prostaglandina, se recomienda la aplicacin de 400 a 700 UI de eCG, realizando un destete temporario de los terneros por 48 hs. En el dcimo da del protocolo se inyecta por va intramuscular 1 mg de Benzoato de Estradiol, realizando la inseminacin artificial a tiempo fijo 24 hs despus. 2.5.2 Protocolos con prostaglandinas.
Doble aplicacin de prostaglandinas en la totalidad de los animales El mtodo tradicional de utilizacin de las prostaglandinas con el objetivo de sincronizacin de celos, prevee la utilizacin de dos dosis de hormona aplicada con un intervalo de 12 a 14 das. La primera aplicacin en rodeos cclicos normalmente el efecto luteoltico se da aproximadamente en el 60% de las vacas. Con la segunda aplicacin de prostaglandina se introduce en estro a la totalidad de los animales. A partir de las 48 horas de la segunda aplicacin se comienza a detectar celo e inseminar por 2 a 3 das. Doble aplicacin de Prostaglandina con inseminacin despus de la primera y segunda dosis Este mtodo consiste en una variante del procedimiento descrito anteriormente utilizado para inseminar vacas que entran en celo despus de la primera aplicacin de prostaglandina. Los animales son observados despus de la primera aplicacin por doce das. Los que no se detectaron en celo, reciben una segunda dosis de prostaglandina y son inseminados cuando demuestran el celo, que se da la mayora de las veces entre las 48 y 96 horas. A pesar de la economa de la hormona, tiene como desventaja en relacin al mtodo original la observacin de un periodo ms largo de celos. Aplicacin nica de prostaglandina despus de un periodo de observacin de celos
Este protocolo se basa en la observacin de celos de las vacas en un periodo de 7 das e inseminacin de las verificadas en celo, siendo aplicada al sptimo da una dosis de prostaglandina en todas las vacas que no ciclaron. El periodo de observacin de siete das debe dar tiempo para que todas las vacas en el momento del segundo tratamiento se encuentren en diestro. Todos los protocolos con prostaglandinas solamente son indicados para animales cclicos, resultando en completo fracaso cuando lo aplicamos en animales con condiciones nutricionales deficitarias y en estado de acclica. Resultados y discusin Conclusin.
Literatura citada. Gadella BM,Tsai P, Boerke A, Brewis IA. Sperm head membrane reorganization during capacitation. Dev Biol. 52: 473. 2008. Inaba K. Molecular architecture of the sperm agella: molecules for motility and signaling. Zool Sci. 20: 1043. 2003. Tulsiani DRP. Glycan-modifying enzymes in luminal uid of the mammalian epididymis: An overview of their potential role in sperm maturation. Mol & Cell Endocr 250: 58. 2006. Flesch FM, Gadella BM. Dynamics of the mammalian sperm plasma membrane in the process of fertilization. Biochim Biophys Acta. 1469: 197. 2000. Cortvrindt, R. & Smitz, J. In vitro follicle growth: achievements in mammalian species. Reprod Domest Anim, v.36, p.3-9, 2001. Hsueh, A.J.W.; Billing, H.; Tsafriri, A. Ovarian follicle atresia: a hormonally controlled apoptotic process. Endocr Rev, 15(6):707-725, 1994. Roberts, S.J. Veterinary obstetrics and genital diseases. Second edition. Ithaca-NY: Ed. Edwars brothers inc, 1971. p.343-375. Fortune, J.E. Activation of primordial follicles. In: Eppig J., Hegele-Hartung C.H., Lessl M. (eds.). The future of the oocyte basic and clinical aspects. Nueva York: Springer, 2002. p.11-21. Ginther, O.J. Selection of the dominant follicle in cattle and horses. Animal Reproduction Science, v.60-61, p.61-79, 2000. Faes, R.F.; Caldas-Bussiere, M.C.; Mosa-E-Silva, A.A.M. et al. Relationship among nitric oxide, progesterone and estradiol-17 concentrations in follicular fluid during follicular development in cattle. Anim reprod, v.4, n.1/2, p.31-37, 2007.
Basini, G.; Baratta, M.; Ponderato, N. et al. Is nitric oxide an autocrine modulator of bovine granulosa cell function? Reprod Fertil Dev, v,10, n.6, p.471-478, 1998. Jolly, P.D.; Smith, P.R; Heath, D.A. et al. Morphological evidence of apoptosis and prevalence of apoptotic versus mitotic cells in membrane granulosa of ovarian follicles during spontaneous and induced atresia in ewes. Biol Reprod, 56:837-846, 1997. Witt, A.A.C. & Kruip, T.A.M. Bovine cumulus oocyte- complex quality is reflected in sensitivity for amanitin, oocyte-diameter and development capacity. Animal Reproduction Science, v.65, p.5165, 2001. Ginther, O.J.; Knopf, L. & Kastelic, J.P. Temporal associations among ovarian events in cattle during oestrous cycles with two and three follicular waves. J. Reprod. Fertil., v.87, p.223-230, 1989a. Adams, G.P.; Matteri, R.L.; Kastelic, J.P. et al. Association between surges of follicle stimulating hormone and the emergence of follicular waves in heifers. J. Reprod. Fert., v.94, n.1, p.177-188, 1992. B, G.A.; Adams, G.P.; Pierson, R.A. Follicular waves dinamic after estradiol 17 treatment of heifers with or without a progesterone implant. Theriogenology, v. 41, p.1555-1569, 1994. Wiltbank, M.C.; Pursley, J.R.; Fricke, P.M. et al. Development of AI and ET programs that do not require detection of estrus using recent information on follicular growth. Proceedings of the XV annual convention AETA, memorias, 1996. 62p. Rajakoski, E. The ovarian follicular system in sexually mature heifers. Acta endocrinolgica, v,34, Suppl. 52, p.1-68, 1960. Binelli, M. Estratgias anti-luteolticas para a melhora da sobrevivnvia embrionria em bovinos. In: Madureira, E.H.; Baruselli, P.S. Controle farmacolgico do ciclo estral em ruminantes. So Paulo: FUNVET, 2000. p.99- 114. Borges, .M.; Torres, C.A.A.; Ruas, J.R.M. et al. Dinmica folicular ovariana em novilhas mestias Holands-Zebu. Arq. Bras.Med.Vet. Zootec., v.53, n.5, p.595-604, 2001. Pierson, R.A. & Ginther, O.J. Ultrasonographic apearence of the bovine uterus during the estrous cycle. J. Vet. Med. Assoc., 190:995- 1002, 1987. Savio, J.D.; Keenan, L.; Boland, M.P. et al. Pattern of growth of dominant follicles during oestrus cycle in heifers. Journal of Reproduction and Fertility, v.83, p.663-671, 1988. Siriois, J. & Fortune, J.E. Ovarian follicular dynamics during the oestrus cycle in heifers monitored by real time ultrasonography. Biology of Reproduction, v.39, p.308-317, 1988. Figueiredo, R.A.; Barros, C.M.; Pinheiro, O.L. et al. Ovarian follicular dynamics in Nelore Breed (Bos indicus). Theriogenology, v.47, p.1489- 505, 1997.
Pinheiro, O.L.; Barros, C.M.; Figueiredo, R.A. et al. Estrus behavior and the estrus-to- ovulation interval in Nelore cattle (Bos indicus) with natural estrus or estrus induced with prostaglandina F2 or norgestomet and estradiol valerate. Theriogenology, v.49, p.667-681, 1998. Baruselli, P.S.; Gimenes, L.U.; Sales, J.N.S. Fisiologia reprodutiva de fmeas taurinas e zebunas. Revista Brasileira de Reproduo Animal, v. 31, n.2, p.205-211, 2007. Rhodes, F.M.; Fitzpatrick, L.A.; Entwistle, K.W. et al. Sequential changes in ovarian follicular dynamics in Bos indicus heifers before and after nutricional anoestrus. Journal of Reproduction and Fertility, v.104, p.41-49, 1995. Viana, J.H.M.; Ferreira, A.M.; S, W.F. et al. Caractersticas morfolgicas e funcionais do corpo lteo durante o ciclo estral em vacas da raa Gir. Arquivo brasileiro de Medicina Veterinria e Zootecnia, v.51, n.3, p.251-256, 1999. Luiz, E.R. 2002. Dinmica Folicular em Bovinos. Monografia. UNESP-Botucatu. Wolfenson, D.; Inbara, G.; Rotha, Z. et al. Follicular dynamics and concentrations of steroids and gonadotropins in lactating cows and nulliparous heifers. Theriogenology, v.62, p.1042-1055, 2004. Carvalho, J.B.; Carvalho, N.A.; Reis, E.L. et al. Effect 97Motta Delgado et al. vet.zootec. 5(2): 88-99, 2011 of early luteolysis in progesterone-based timed AI protocols in Bos indicus, Bos indicus x Bos taurus, and Bos taurus heifers. Theriogenology, v.69, n.2, p.167-175, 2008. Ginther O.J.; Wiltbank, M.C.; Fricke, P.M. et al. Selection of the dominant follicle in cattle. Biol Reprod, v.55, p.1187-1194, 1996. Silcoux, R.W.; Powell, K.L.; Kiser, T.E. Ability of dominant follicles to respond to exogenous GnRH administration is dependent on their stage of development. Journal of Animal Science, v.71, suppl. 1, p.219 (Abstract), 1993. Murphy, M.G.; Boland, M.P.; Roche, J.F. Pattern of follicular growth and resumption of ovarian activity in post partum beff suckler cows. Journal and Reproduction and Fertility, v.90, p.523-533, 1990. Badinga, L.; Thatcher, W.W.; Wilcox, C.J. et al. Effect of season on follicular dynamics and plasma concentrations of estradiol-17b, progesterone and luteinizing hormone in lactating Holstein cows. Theriogenology, v.42, n.8, p.1263- 1274, 1994. Richards, MW, JC Spitzer, y MB Warner. 1986 . Efecto de la variacin de los niveles de nutricin posparto y condicin corporal al parto sobre el comportamiento reproductivo posterior en el ganado vacuno. J. Anim. . Sci.62 : 300 -306. Kirby, CJ, MF Smith, DH Keisler y MC Lucy. 1997. Funcin folicular en vacas lecheras tratadas con liberacin sostenida somatotropina bovina. J.Lcteos. . Sci. 80: 273 -285.
Kirby, CJ, MF Smith, DH Keisler y MC Lucy. 1997. Funcin folicular en vacas lecheras tratadas con liberacin sostenida somatotropina bovina. J.Lcteos. . Sci. 80: 273 -285. Perry, GA, MF Smith y DJ Patterson. 2002. Evaluacin de un tiempo fijoartificial inseminacin posparto protocolo para vacas de carne amamantado. J. Anim. . Sci. 80: 3060 -3064. Stegner, JE, JF Bader, FN Kojima, MR Ellersieck, MF Smith y DJ Patterson.2004a. Fijo a tiempo artificial inseminacin de vacas de cra a las 72 80 horas despus del tratamiento con el MGA Seleccionar protocolo.Theriogenology 61: 1299 -1305. Kojima, FN, JE Stegner, BE Salfen, RL Eakins, MF Smith y DJ Patterson. 2002. Un programa de inseminacin artificial a tiempo fijo en vacas de carne con 7-11 Synch. Proc. West. Sec. Am. Soc. Anim. . Sci. 53: 411 -413. Kojima, FN, JE Stegner, JF Bader, DJ Schafer, RL Eakins, MF Smith y DJ Patterson. 2003a. Un programa de inseminacin artificial a tiempo fijo con 7-11 Synch. Proc. West. Sec. Am. Soc. Anim. . Sci. 54: 265 -267. Kojima, FN, BE Salfen, WA Ricke, MC Lucy, MF Smith y DJ Patterson. 2000 .Desarrollo de un protocolo de sincronizacin de estro para el ganado vacuno con alimentacin a corto plazo de acetato de melengestrol:. 7-11 Synch J.Anim. . Sci. 78: 2186 -2191. Patterson, DJ, JE Stegner, FN Kojima y MF Smith. 2002. MGA Select mejora la respuesta del estro en vacas de cra en situaciones acompaadas con altas tasas de anestro. Proc. West. Sec. Am. Soc. Anim. . Sci. 53: 418 -420. Stegner, JE, FN Kojima, MR Ellersieck, MC Lucy, MF Smith y DJ Patterson.2004b. Una comparacin de los protocolos basados en progestina para sincronizar el estro posparto en vacas de carne. J. Anim. . Sci. 82: 1016-1021. Perry, GA 2003. La induccin del estado de ciclismo y efecto del tamao del folculo sobre la fertilidad en vacas de carne posparto. Ph.D. Diss., Univ.de Missouri, Columbia. Vasconcelos, JL, R. Sartori, HN Oliveira, JG Guenther y MC Wiltbank. 2001 .Reduccin del tamao del folculo ovulatorio reduce el subsecuente tamao de ltea y la tasa de embarazo. Theriogenology 56: 307 -314.
Stegner, JE, FN Kojima, MR Ellersieck, MC Lucy, MF Smith y DJ Patterson. 2004c. Dinmica folicular y perfiles de esteroides en las vacas durante y despus del tratamiento con protocolos basados en progestina para la sincronizacin del estro. J. Anim. . Sci. 82 : 1022 -1028. Kojima, FN, JF Bader, JE Stegner, MF Smith y DJ Patterson. 2003b. Una comparacin de dos programas de IA a tiempo fijo para vacas de cra. J. Anim.. Sci. 81 (Supl. 1): 50. (Abstr.).
Mussard, ML, CR Burke, CL Gasser, EJ Behlke, KA Colliflower, DE Grum, y el Da de ML. 2003. Respuesta ovulatoria, la funcin ltea y fertilidad en el ganado inducidas a ovular folculos dominantes de maduracin temprana o tarda. Biol. Reprod. 68 (Supl. 1): 332. (Abstr.).
Das könnte Ihnen auch gefallen
- Tesis Doctoral: Universidad de Granada Programa de Bioquímica y Biología MolecularDokument165 SeitenTesis Doctoral: Universidad de Granada Programa de Bioquímica y Biología MolecularNicole LlanosNoch keine Bewertungen
- 2 Clase - HormonasDokument55 Seiten2 Clase - HormonasIcaro Valentin FariaNoch keine Bewertungen
- YeguaDokument37 SeitenYeguaLuisa QuirogaNoch keine Bewertungen
- Cuál Es La Función de Túnica AlbugíneaDokument2 SeitenCuál Es La Función de Túnica AlbugíneaPC Tzul100% (1)
- Gineco CEARDokument254 SeitenGineco CEARPANDA- ImpresionesNoch keine Bewertungen
- Informe 2 de ReproDokument6 SeitenInforme 2 de ReproGloria Torres ChumbeNoch keine Bewertungen
- Interacción de GametosDokument17 SeitenInteracción de GametosYolotzin CoronaNoch keine Bewertungen
- Anatomia Del Aparato Reproductor Femenino-1Dokument76 SeitenAnatomia Del Aparato Reproductor Femenino-1Ana Laura JerezNoch keine Bewertungen
- Fisiología Eje y Ciclo Sexual FemeninoDokument30 SeitenFisiología Eje y Ciclo Sexual FemeninoJUAN CARLOS GARCIA CRUZNoch keine Bewertungen
- Resumen de OntogeniaDokument23 SeitenResumen de OntogeniaJatnielNoch keine Bewertungen
- Trabajo de Anatomía Ciclo MenstrualDokument27 SeitenTrabajo de Anatomía Ciclo MenstrualCristian David Hurtado PeñaNoch keine Bewertungen
- RESUMEN GINECOLOGIA de Repaso 2018.docx Versión 1Dokument45 SeitenRESUMEN GINECOLOGIA de Repaso 2018.docx Versión 1bautista romanNoch keine Bewertungen
- Resumen GineDokument15 SeitenResumen GineNicol Rojas AstorgaNoch keine Bewertungen
- Fisiologia Eje Hipotalamo Hipofifi OvarioDokument110 SeitenFisiologia Eje Hipotalamo Hipofifi OvarioJunior Ponce Melendez100% (1)
- Resumen Etapa PrenatalDokument26 SeitenResumen Etapa PrenatalWaldo Lastra MoralesNoch keine Bewertungen
- Ciclo OváricoDokument12 SeitenCiclo OváricoHeidy LozadaNoch keine Bewertungen
- Ciclo MenstrualDokument33 SeitenCiclo Menstrualsotelorosa568Noch keine Bewertungen
- Articulo Ciclo MenstrualDokument16 SeitenArticulo Ciclo MenstrualDANIELA MELISSA GARRIDO VÁSQUEZNoch keine Bewertungen
- 238 Determinación de La Viabilidad de Ovocitos Bovinos Obtenidos Post Mortem A Varios Periodos de Tiempo en El Camal MunicipalDokument75 Seiten238 Determinación de La Viabilidad de Ovocitos Bovinos Obtenidos Post Mortem A Varios Periodos de Tiempo en El Camal MunicipaljuanNoch keine Bewertungen
- Manual de Reproduccion Bovina - EmbriologiaDokument16 SeitenManual de Reproduccion Bovina - EmbriologiaAndyssyta KesstaNoch keine Bewertungen
- Gametogénesis 2022Dokument53 SeitenGametogénesis 2022Valen a pNoch keine Bewertungen
- 186-Reprod CompendioDokument144 Seiten186-Reprod CompendioRita Cardozo100% (1)
- Resumen Salud Sexual y ReproductivaDokument44 SeitenResumen Salud Sexual y ReproductivaDiscopy Dis CopyNoch keine Bewertungen
- Hemorragia Del III Trimestre, CICLO MENSTRUALDokument11 SeitenHemorragia Del III Trimestre, CICLO MENSTRUALAmram Francisco Revilla GonzalezNoch keine Bewertungen
- Micro Resumen Ciclo Ovarico y EndometrialDokument4 SeitenMicro Resumen Ciclo Ovarico y EndometrialCriisthii HernandezNoch keine Bewertungen
- Ciclo Estral en BovinosDokument36 SeitenCiclo Estral en BovinosJorge VergaraNoch keine Bewertungen
- Lic. Marcos Fabio Helguero Enfermería Materno InfantilDokument628 SeitenLic. Marcos Fabio Helguero Enfermería Materno InfantilBruno ZverdaNoch keine Bewertungen
- Ciclo Ovarico y Seg FolicularDokument41 SeitenCiclo Ovarico y Seg FolicularPierre GonzalesNoch keine Bewertungen
- Wuolah Free Embrio Notas JuntasDokument256 SeitenWuolah Free Embrio Notas JuntasRafael DavidNoch keine Bewertungen
- Fisiología de La MenstruaciónDokument24 SeitenFisiología de La MenstruaciónMariana MataNoch keine Bewertungen