Das könnte Ihnen auch gefallen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
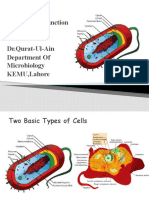
- Bacterial Cell Structure FunctionDokument35 SeitenBacterial Cell Structure FunctionItzz MilzzNoch keine Bewertungen
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- Microbiology, Usmle EndpointDokument208 SeitenMicrobiology, Usmle EndpointYazan M Abu-FaraNoch keine Bewertungen
- Bacterial Cell Structure 1) Capsule: Spirillum (Spiral) - Mycoplasmas Are BacteriaDokument4 SeitenBacterial Cell Structure 1) Capsule: Spirillum (Spiral) - Mycoplasmas Are BacteriaTLCRNoch keine Bewertungen
- Regulat® CompendiumDokument90 SeitenRegulat® CompendiumBiologicalMed99100% (3)
- Medical Micro-Apmc QuestionsDokument8 SeitenMedical Micro-Apmc QuestionsDonnaBells Hermo LabaniegoNoch keine Bewertungen
- Cholera - WhoDokument5 SeitenCholera - WhoSuci D PutriNoch keine Bewertungen
- Cholera - CDCDokument14 SeitenCholera - CDCSuci D PutriNoch keine Bewertungen
- Breast Anatomy Phisiology NormalDokument11 SeitenBreast Anatomy Phisiology NormalSuci D PutriNoch keine Bewertungen
- Benign Breast DiseaseDokument9 SeitenBenign Breast DiseaseSuci D PutriNoch keine Bewertungen
- SalmonellosisDokument40 SeitenSalmonellosispurposef49Noch keine Bewertungen
- Koch's Postulates and the Chain of InfectionDokument7 SeitenKoch's Postulates and the Chain of InfectionHERRERA, ANGELANoch keine Bewertungen
- Characteristic of Enteric BacteriaDokument56 SeitenCharacteristic of Enteric BacteriaAlpesh SutharNoch keine Bewertungen
- Immunology (Levinson)Dokument93 SeitenImmunology (Levinson)ECFMGNoch keine Bewertungen
- Article1407502547 - Shale Et AlDokument14 SeitenArticle1407502547 - Shale Et AlGoummeli6 SocratesNoch keine Bewertungen
- Lab 3 Bacterial Staining Techniques IIDokument10 SeitenLab 3 Bacterial Staining Techniques IIErreneoNoch keine Bewertungen
- Coup de ChaleurDokument16 SeitenCoup de ChaleurEric CharpentierNoch keine Bewertungen
- Leaky Gut GuideDokument6 SeitenLeaky Gut GuideKrishna DasNoch keine Bewertungen
- Mcrobio Cell MembraneDokument8 SeitenMcrobio Cell MembraneResty De Guzman SoteloNoch keine Bewertungen
- Fever and Fever of Unknown Origin: Review, Recent Advances, and Lingering DogmaDokument12 SeitenFever and Fever of Unknown Origin: Review, Recent Advances, and Lingering DogmantnquynhproNoch keine Bewertungen
- USMLE Step 1 First Aid 2020 30th - MICROBIOLOGYDokument82 SeitenUSMLE Step 1 First Aid 2020 30th - MICROBIOLOGYArt PuffNoch keine Bewertungen
- 69 Haines Ely-Is Psoriasis A Bowel DiseaseDokument14 Seiten69 Haines Ely-Is Psoriasis A Bowel DiseaseJoe DoeNoch keine Bewertungen
- Biology of Nutrition in Growing AnimalsDokument599 SeitenBiology of Nutrition in Growing AnimalsSam Rafael Bravo GutierrezNoch keine Bewertungen
- CarbohydratDokument6 SeitenCarbohydratVan Anh NguyenNoch keine Bewertungen
- Chapter 10 Immunity and InfectionDokument96 SeitenChapter 10 Immunity and InfectionTofikNoch keine Bewertungen
- 85 - Bacterial Endotoxin Test - Usp MonographDokument10 Seiten85 - Bacterial Endotoxin Test - Usp MonographVanessa CarreraNoch keine Bewertungen
- Bacillus lipopeptides: versatile weapons against plant diseasesDokument11 SeitenBacillus lipopeptides: versatile weapons against plant diseasesjazminNoch keine Bewertungen
- p17512c Guia de Pellicon MaintenanceDokument20 Seitenp17512c Guia de Pellicon MaintenanceEdgar Ortiz AhumadaNoch keine Bewertungen
- Lesson 28Dokument12 SeitenLesson 28Sheikh FaishalNoch keine Bewertungen
- Metformin Retards Aging in C. Elegans by Altering Microbial Folate and Methionine MetabolismDokument12 SeitenMetformin Retards Aging in C. Elegans by Altering Microbial Folate and Methionine MetabolismAndrew CitrawanNoch keine Bewertungen
- Mtap2 Bacte 1Dokument27 SeitenMtap2 Bacte 1DENISE MARA�ANoch keine Bewertungen
- Biofilms Microbial DONLAN 2002Dokument138 SeitenBiofilms Microbial DONLAN 2002Anushka Sujeewon100% (1)
- Package Insert - PYROSPERSE™ Dispersing Agent (English) - Original - 27876Dokument7 SeitenPackage Insert - PYROSPERSE™ Dispersing Agent (English) - Original - 27876Putri DozanNoch keine Bewertungen
- Microbiology - Bacte, Myco, ViroDokument120 SeitenMicrobiology - Bacte, Myco, ViroSogan, MaureenNoch keine Bewertungen
- Guideline For Dialysate Quality of Spanish Society of Nephrology (Second Edition, 2015)Dokument52 SeitenGuideline For Dialysate Quality of Spanish Society of Nephrology (Second Edition, 2015)luc1902Noch keine Bewertungen