Das könnte Ihnen auch gefallen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (120)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Enneagram Type-3Dokument17 SeitenEnneagram Type-3pundir100% (1)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- The Fanes of BaroviaDokument8 SeitenThe Fanes of BaroviaFelipe100% (1)
- Out of The Trap Selected Lectures of Alan W. Watts PDFDokument164 SeitenOut of The Trap Selected Lectures of Alan W. Watts PDFAKU95100100% (2)
- I AM AMERICA 5 Golden City Vortex Safe Zones 9-2009updaterDokument14 SeitenI AM AMERICA 5 Golden City Vortex Safe Zones 9-2009updaterDaniel HastayNoch keine Bewertungen
- Underground Installation of Thermoplastic Pipe For Sewers and Other Gravity-Flow ApplicationsDokument10 SeitenUnderground Installation of Thermoplastic Pipe For Sewers and Other Gravity-Flow ApplicationsDavid SolisNoch keine Bewertungen
- Corrosion and PaintingDokument13 SeitenCorrosion and PaintingManeet GogiaNoch keine Bewertungen
- Sandstone FloraDokument10 SeitenSandstone FloraAnon BoletusNoch keine Bewertungen
- PPC TextbookDokument289 SeitenPPC Textbookjaywan91Noch keine Bewertungen
- Dissonant Heritage. The Management of The Past As A Resource in Conflict. J.E. Tunbridge and G.J. AshworthDokument305 SeitenDissonant Heritage. The Management of The Past As A Resource in Conflict. J.E. Tunbridge and G.J. AshworthFRANCISCO ALBERTO DE LA ROSA MANJARREZ100% (1)
- Chemical Resistance Chart GlovesDokument8 SeitenChemical Resistance Chart Glovesjitendertalwar1603Noch keine Bewertungen
- Steam TableDokument6 SeitenSteam Tablexian_putraNoch keine Bewertungen
- Building Code InfoDokument10 SeitenBuilding Code InfoDewi GlorianaNoch keine Bewertungen
- Contractors All Risks InsuranceDokument2 SeitenContractors All Risks InsuranceDewi GlorianaNoch keine Bewertungen
- Nfpa82 2004 EditionDokument3 SeitenNfpa82 2004 EditionDewi GlorianaNoch keine Bewertungen
- Building Heating SystemsDokument28 SeitenBuilding Heating SystemsDewi GlorianaNoch keine Bewertungen
- South Milwaukee RedactedDokument16 SeitenSouth Milwaukee RedactedDewi GlorianaNoch keine Bewertungen
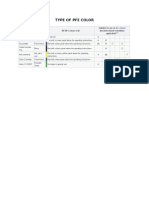
- Type of Pfe ColorDokument1 SeiteType of Pfe ColorDewi GlorianaNoch keine Bewertungen
- Fire and Rescue Protection OrdinanceDokument36 SeitenFire and Rescue Protection OrdinanceDewi GlorianaNoch keine Bewertungen
- 2010-12-29 Standards of Construction For Health Care FacilitiesDokument220 Seiten2010-12-29 Standards of Construction For Health Care FacilitiesDaniel BellemareNoch keine Bewertungen
- NFPA Fire DefenderDokument1 SeiteNFPA Fire DefenderDewi GlorianaNoch keine Bewertungen
- Industry-Farmer Alliance-Need of The HourDokument2 SeitenIndustry-Farmer Alliance-Need of The HourDewi GlorianaNoch keine Bewertungen
- Chemical Engineering January 2013Dokument68 SeitenChemical Engineering January 2013Dewi GlorianaNoch keine Bewertungen
- 40 Inventive Principle - Chemical Enginering ExamplesDokument14 Seiten40 Inventive Principle - Chemical Enginering ExamplesDewi GlorianaNoch keine Bewertungen
- ComputationDokument27 SeitenComputationDewi GlorianaNoch keine Bewertungen
- CH 1 Intro To LeanDokument26 SeitenCH 1 Intro To LeanIslam FouadNoch keine Bewertungen
- Corrosion1 0Dokument47 SeitenCorrosion1 0Akshat GroverNoch keine Bewertungen
- Biology ConclusionDokument2 SeitenBiology Conclusionapi-297841438Noch keine Bewertungen
- CH 7 Assignment DPS 2Dokument2 SeitenCH 7 Assignment DPS 2V BNoch keine Bewertungen
- End of Term Test Exercise 1: Read The Following Text and Decide Whether The Following Statements Are TrueDokument3 SeitenEnd of Term Test Exercise 1: Read The Following Text and Decide Whether The Following Statements Are TrueHương ChanelNoch keine Bewertungen
- Bustamante Becerra Bitencourt2007Dokument16 SeitenBustamante Becerra Bitencourt2007Sandro Begazo RodriguezNoch keine Bewertungen
- Norma ASTM A380.27738-1Dokument12 SeitenNorma ASTM A380.27738-1windeletaNoch keine Bewertungen
- Liveable Neighbour Hoods Community Design Code Ed 1 - GWA Australia - 1997Dokument4 SeitenLiveable Neighbour Hoods Community Design Code Ed 1 - GWA Australia - 1997PublicHealthbyDesignNoch keine Bewertungen
- Il Piano Ecoturistico Del FayoumDokument160 SeitenIl Piano Ecoturistico Del FayoumayepatagonikNoch keine Bewertungen
- Memorandum: Project: Library Archives Museum Project Date: 11 30 2010 Subject: Alternate Sites Feasibility StudyDokument7 SeitenMemorandum: Project: Library Archives Museum Project Date: 11 30 2010 Subject: Alternate Sites Feasibility StudyECI HyerNoch keine Bewertungen
- Wildlife in ChhattisgarhDokument12 SeitenWildlife in ChhattisgarhAbhinay D Raj Abhinay D RajNoch keine Bewertungen
- Deep Water by William DouglasDokument10 SeitenDeep Water by William DouglasAyu HarshalNoch keine Bewertungen
- Sumatra - Geology, Resources and TectonicsDokument35 SeitenSumatra - Geology, Resources and Tectonicsadinda novianaNoch keine Bewertungen
- George Kelly: 2) Individuality CorollaryDokument2 SeitenGeorge Kelly: 2) Individuality CorollaryJeffrey ManzanoNoch keine Bewertungen
- Tunneling in HimalayasDokument4 SeitenTunneling in HimalayassaritasohamNoch keine Bewertungen
- My PersonalityDokument2 SeitenMy Personalityapi-3710687400% (1)
- English Vistas Chapter 2 The Tiger KingDokument7 SeitenEnglish Vistas Chapter 2 The Tiger Kingshamiksha dNoch keine Bewertungen
- A Mineral System ApproachDokument4 SeitenA Mineral System ApproachKaharuddin HawingNoch keine Bewertungen
- Level 9 Passage 4Dokument3 SeitenLevel 9 Passage 4JefaradocsNoch keine Bewertungen
- VolcanoesDokument32 SeitenVolcanoesapi-302968888Noch keine Bewertungen
- Acid ResistanceDokument2 SeitenAcid ResistanceBatepola BacNoch keine Bewertungen
- ENTR 20113 - Chapter 1Dokument6 SeitenENTR 20113 - Chapter 1Hanny ValenciaNoch keine Bewertungen