Das könnte Ihnen auch gefallen
- U4 - Estimación Por Intervalos de Confianza y Tipos de MuestreoDokument64 SeitenU4 - Estimación Por Intervalos de Confianza y Tipos de MuestreoAlondraNoch keine Bewertungen
- Cura Herida TalónDokument1 SeiteCura Herida TalónFlavia González LimaNoch keine Bewertungen
- 6 Mama y Mediastino TextoDokument3 Seiten6 Mama y Mediastino TextoFlavia González LimaNoch keine Bewertungen
- 3 Retrosoma - TextoDokument3 Seiten3 Retrosoma - TextoFlavia González LimaNoch keine Bewertungen
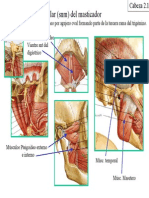
- 2 Cabeza - 2aDokument3 Seiten2 Cabeza - 2aFlavia González LimaNoch keine Bewertungen
- 5 Pared Toracica y Abdominal TextoDokument3 Seiten5 Pared Toracica y Abdominal TextoFlavia González LimaNoch keine Bewertungen
- 2 Anatomia de La Cabeza IIDokument5 Seiten2 Anatomia de La Cabeza IIFlavia González LimaNoch keine Bewertungen
- 4 Extremidades - TextoDokument3 Seiten4 Extremidades - TextoFlavia González LimaNoch keine Bewertungen
- Estructura y componentes de la mama y el mediastinoDokument4 SeitenEstructura y componentes de la mama y el mediastinoFlavia González LimaNoch keine Bewertungen
- 5 Pared Del Torax y Abdomen ImagenesDokument4 Seiten5 Pared Del Torax y Abdomen ImagenesFlavia González LimaNoch keine Bewertungen
- 4 Extremidades ImagenesDokument5 Seiten4 Extremidades ImagenesFlavia González LimaNoch keine Bewertungen
- 3 Retrosoma ImagenesDokument11 Seiten3 Retrosoma ImagenesFlavia González LimaNoch keine Bewertungen
- 3 Retrosoma ImagenesDokument11 Seiten3 Retrosoma ImagenesFlavia González LimaNoch keine Bewertungen
- 1 Anatomia de La Cabeza 1Dokument3 Seiten1 Anatomia de La Cabeza 1Flavia González LimaNoch keine Bewertungen
- 1 A+imágenDokument5 Seiten1 A+imágenFlavia González LimaNoch keine Bewertungen
- .. DocsUp 244 131Dokument3 Seiten.. DocsUp 244 131Flavia González LimaNoch keine Bewertungen
- 2008 Junio PDFDokument2 Seiten2008 Junio PDFJorge BáezNoch keine Bewertungen
- GLÚCIDOSDokument10 SeitenGLÚCIDOSFlavia González LimaNoch keine Bewertungen
- Intestino Delgado y Grueso Enfer 07-08Dokument4 SeitenIntestino Delgado y Grueso Enfer 07-08Flavia González LimaNoch keine Bewertungen
- BacteriasDokument10 SeitenBacteriasFlavia González LimaNoch keine Bewertungen
- Hi Alo PlasmaDokument11 SeitenHi Alo PlasmaFlavia González LimaNoch keine Bewertungen
- Problemas de Genética Aprobados El 3-4-03Dokument2 SeitenProblemas de Genética Aprobados El 3-4-03Flavia González LimaNoch keine Bewertungen
- LÍPIDOSDokument8 SeitenLÍPIDOSFlavia González LimaNoch keine Bewertungen
- VirusDokument11 SeitenVirusFlavia González LimaNoch keine Bewertungen
- INMUNODokument22 SeitenINMUNOFlavia González LimaNoch keine Bewertungen
- Por Tierras de EspañaDokument4 SeitenPor Tierras de EspañaFlavia González LimaNoch keine Bewertungen
- Tema 16 Transición y DemocraciaDokument12 SeitenTema 16 Transición y DemocraciaFlavia González LimaNoch keine Bewertungen
- PROTEÍNASDokument12 SeitenPROTEÍNASFlavia González LimaNoch keine Bewertungen
- Transformaciones Económicas y Sociales en El Primer Tercio Del Siglo XXDokument7 SeitenTransformaciones Económicas y Sociales en El Primer Tercio Del Siglo XXFlavia González LimaNoch keine Bewertungen
- Textos Historia de EspañaDokument15 SeitenTextos Historia de EspañaConcha Lopez GutierrezNoch keine Bewertungen
- Abordaje Endoaural ModificadoDokument6 SeitenAbordaje Endoaural ModificadoRaul MartellNoch keine Bewertungen
- Prueba DetalleDokument1 SeitePrueba DetalleMARIA ALEJANDRA GUILLEN ZAMBRANONoch keine Bewertungen
- Informe 001 de La Planta AurelsaDokument19 SeitenInforme 001 de La Planta AurelsaMilagros Allcca NoaNoch keine Bewertungen
- Documento Básico de Contratación de Proyectos en BoliviaDokument31 SeitenDocumento Básico de Contratación de Proyectos en BoliviaRamiro Cruz VillarroelNoch keine Bewertungen
- Mascarillas Caseras para El CabelloDokument9 SeitenMascarillas Caseras para El CabelloGiselaSauceNoch keine Bewertungen
- Ejercicio # 1lexico MedicoDokument2 SeitenEjercicio # 1lexico MedicoHenry Leroy Lewis BatresNoch keine Bewertungen
- MédicoDokument5 SeitenMédicoIvanky Vander WoodsenNoch keine Bewertungen
- Resumen de Metrados Arq - SLDokument4 SeitenResumen de Metrados Arq - SLJimmy Heisen Julca VillanuevaNoch keine Bewertungen
- Diagnóstico educativo integral 2015-2016Dokument6 SeitenDiagnóstico educativo integral 2015-2016jhonny265Noch keine Bewertungen
- Presentacion de Bienestar NuxgenDokument46 SeitenPresentacion de Bienestar NuxgenJorge Luis Martinez RodriguezNoch keine Bewertungen
- 18649-Texto Del Artículo-73935-1-10-20170602Dokument11 Seiten18649-Texto Del Artículo-73935-1-10-20170602Valentina CastañedaNoch keine Bewertungen
- La Entrevista MotivacionalDokument11 SeitenLa Entrevista Motivacionalapi-26645842100% (1)
- Examen Parcial Teoria de Sistemas - ClaveDokument5 SeitenExamen Parcial Teoria de Sistemas - Clavelibrada moralesNoch keine Bewertungen
- Portafolio 1er Parcial - Melanie Garcia ReyesDokument53 SeitenPortafolio 1er Parcial - Melanie Garcia ReyesMelanie GarciaNoch keine Bewertungen
- Control sísmica mesaDokument159 SeitenControl sísmica mesaDiego CamachoNoch keine Bewertungen
- 7 Decisiones (El Regalo Del Viajero)Dokument8 Seiten7 Decisiones (El Regalo Del Viajero)AlejandroB.CoaguilaNoch keine Bewertungen
- 12 Pasos de Un Pitch..Dokument28 Seiten12 Pasos de Un Pitch..Milagros Alarcon MoyedaNoch keine Bewertungen
- Evidencia Diagrama Identificar Proceso Preparacion BiofertilizanteDokument2 SeitenEvidencia Diagrama Identificar Proceso Preparacion Biofertilizantefabio romeroNoch keine Bewertungen
- RECUBRIMIENTOSDokument42 SeitenRECUBRIMIENTOSjesusNoch keine Bewertungen
- Contenido Semana2 U1 ADM101Dokument24 SeitenContenido Semana2 U1 ADM101Tablas MixNoch keine Bewertungen
- Actividad 4Dokument8 SeitenActividad 4Luz Angela Henao NavarroNoch keine Bewertungen
- Lego Mindstorms AvanzadoDokument8 SeitenLego Mindstorms AvanzadoSofia100% (1)
- Eda3 1ero Ccss 2023 Prof. Janet OrtegaDokument9 SeitenEda3 1ero Ccss 2023 Prof. Janet OrtegaHenry JacobiNoch keine Bewertungen
- Sopa de LetrasDokument8 SeitenSopa de LetrasluamanuelalondonoNoch keine Bewertungen
- Identificación de Impactos Ambientales y Estrategias de Solución para La Actividad Floricultora en La Empresa Falcon Farms Del Municipio de Suesca, CundinamarcaDokument24 SeitenIdentificación de Impactos Ambientales y Estrategias de Solución para La Actividad Floricultora en La Empresa Falcon Farms Del Municipio de Suesca, CundinamarcaJuanPabloSuarezBautistaNoch keine Bewertungen
- La Edad de Los CambiosDokument12 SeitenLa Edad de Los CambiosKate Hernández.Noch keine Bewertungen
- TIPOSDEGLOBOSPDFDokument6 SeitenTIPOSDEGLOBOSPDFFrancis RodriguezNoch keine Bewertungen
- TARIFA ISOLTUBEX 2022 3aedicionDokument83 SeitenTARIFA ISOLTUBEX 2022 3aedicionSantiago ChauNoch keine Bewertungen
- Gian Pier Garcia Martinez Gian Pier Etp s13Dokument5 SeitenGian Pier Garcia Martinez Gian Pier Etp s13DAVID UMBO FERNANDEZNoch keine Bewertungen