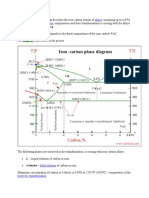
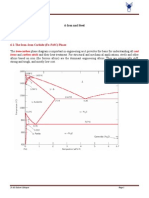
Das könnte Ihnen auch gefallen
- Hatzell Et Al - 2020 - Challenges in Lithium Metal Anodes For Solid-State BatteriesDokument13 SeitenHatzell Et Al - 2020 - Challenges in Lithium Metal Anodes For Solid-State BatteriesJalcamNoch keine Bewertungen
- Hwang Et Al - 2009 - Optimization of Cycloidal Water Turbine and The Performance Improvement byDokument9 SeitenHwang Et Al - 2009 - Optimization of Cycloidal Water Turbine and The Performance Improvement byJalcamNoch keine Bewertungen
- Ubando Et Al - 2022 - Biohydrogen in A Circular BioeconomyDokument16 SeitenUbando Et Al - 2022 - Biohydrogen in A Circular BioeconomyJalcamNoch keine Bewertungen
- Sun - Liu - 2009 - Purification, Structure and Immunobiological Activity of A Water-SolubleDokument4 SeitenSun - Liu - 2009 - Purification, Structure and Immunobiological Activity of A Water-SolubleJalcamNoch keine Bewertungen
- Singh Et Al - 2019 - Biotechnological Applications of Inulin-Rich FeedstocksDokument13 SeitenSingh Et Al - 2019 - Biotechnological Applications of Inulin-Rich FeedstocksJalcamNoch keine Bewertungen
- Ismail Et Al - 2010 - Thermophilic Biohydrogen Production From Palm Oil Mill Effluent (POME) UsingDokument6 SeitenIsmail Et Al - 2010 - Thermophilic Biohydrogen Production From Palm Oil Mill Effluent (POME) UsingJalcamNoch keine Bewertungen
- Kwon - Castaldi - 2008 - Scrap Tires in Waste-to-Energy PlantsDokument2 SeitenKwon - Castaldi - 2008 - Scrap Tires in Waste-to-Energy PlantsJalcamNoch keine Bewertungen
- Zauner Et Al - 2017 - Experimental Characterization and Simulation of A Hybrid Sensible-Latent HeatDokument14 SeitenZauner Et Al - 2017 - Experimental Characterization and Simulation of A Hybrid Sensible-Latent HeatJalcamNoch keine Bewertungen
- Zhang Et Al - 2017 - Life Cycle Assessment of Power-to-GasDokument13 SeitenZhang Et Al - 2017 - Life Cycle Assessment of Power-to-GasJalcamNoch keine Bewertungen
- Pangu - Feke - 2005 - Analysis of Droplet Coalescence in Emulsions Subjected To Acoustic FieldsDokument11 SeitenPangu - Feke - 2005 - Analysis of Droplet Coalescence in Emulsions Subjected To Acoustic FieldsJalcamNoch keine Bewertungen
- Fife - 2007 - Coconut and Its Healing PropertiesDokument6 SeitenFife - 2007 - Coconut and Its Healing PropertiesJalcamNoch keine Bewertungen
- Lourinho, Brito - 2015 - Assessment of Biomass Energy Potential in A Region of Portugal (AltoAlentejo)Dokument13 SeitenLourinho, Brito - 2015 - Assessment of Biomass Energy Potential in A Region of Portugal (AltoAlentejo)JalcamNoch keine Bewertungen
- Savidov Et Al - 2007 - Fish and Plant Production in A Recirculating Aquaponic SystemDokument14 SeitenSavidov Et Al - 2007 - Fish and Plant Production in A Recirculating Aquaponic SystemJalcamNoch keine Bewertungen
- Rotich Et Al - 2009 - Substrate Effects of Grain Spawn Production On Mycelium Growth of OysterDokument4 SeitenRotich Et Al - 2009 - Substrate Effects of Grain Spawn Production On Mycelium Growth of OysterJalcamNoch keine Bewertungen
- Pongprasert Et Al - 2020 - Effect of Carbon Dioxide Micro Bubbles in Combination With Chlorine DioxideDokument6 SeitenPongprasert Et Al - 2020 - Effect of Carbon Dioxide Micro Bubbles in Combination With Chlorine DioxideJalcamNoch keine Bewertungen
- Salomone Et Al - 2023 - Process Simulation and Energy Analysis of Synthetic Natural Gas Production FromDokument17 SeitenSalomone Et Al - 2023 - Process Simulation and Energy Analysis of Synthetic Natural Gas Production FromJalcamNoch keine Bewertungen
- Chen Et Al - 2022 - Design of A Biomass-Heating Network With An Integrated Heat PumpDokument22 SeitenChen Et Al - 2022 - Design of A Biomass-Heating Network With An Integrated Heat PumpJalcamNoch keine Bewertungen
- Kim Et Al - 2020 - Timing and Pattern of Anthocyanin Accumulation During Grain Filling in PurpleDokument7 SeitenKim Et Al - 2020 - Timing and Pattern of Anthocyanin Accumulation During Grain Filling in PurpleJalcamNoch keine Bewertungen
- Orsini Et Al - 2010 - Simplified Hydroponic Floating Systems For Vegetable Production in TrujilloDokument6 SeitenOrsini Et Al - 2010 - Simplified Hydroponic Floating Systems For Vegetable Production in TrujilloJalcamNoch keine Bewertungen
- Kantola, Saari - 2013 - Renewable vs. Traditional Energy Management Solutions - A Finnish Hospital Facility CaseDokument7 SeitenKantola, Saari - 2013 - Renewable vs. Traditional Energy Management Solutions - A Finnish Hospital Facility CaseJalcamNoch keine Bewertungen
- Arafat, Jijakli, Ahsan - 2013 - Environmental Performance and Energy Recovery Potential of Five Processes For Municipal Solid Waste TreaDokument8 SeitenArafat, Jijakli, Ahsan - 2013 - Environmental Performance and Energy Recovery Potential of Five Processes For Municipal Solid Waste TreaJalcamNoch keine Bewertungen
- Zhao Et Al - 2010 - Two Calculation Methods For Greenhouse Gas Emissions From Municipal Solid WasteDokument8 SeitenZhao Et Al - 2010 - Two Calculation Methods For Greenhouse Gas Emissions From Municipal Solid WasteJalcamNoch keine Bewertungen
- Varanda, Pinto, Martins - 2011 - Life Cycle Analysis of Biodiesel ProductionDokument8 SeitenVaranda, Pinto, Martins - 2011 - Life Cycle Analysis of Biodiesel ProductionJalcamNoch keine Bewertungen
- Zahariev, Penkov, Aladjadjiyan - 2014 - Biogas From Animal Manure - Perspectives and Barriers in BulgariaDokument11 SeitenZahariev, Penkov, Aladjadjiyan - 2014 - Biogas From Animal Manure - Perspectives and Barriers in BulgariaJalcamNoch keine Bewertungen
- Flow Chart: Fresh Single Strength Juice (Red or White) With SO2Dokument2 SeitenFlow Chart: Fresh Single Strength Juice (Red or White) With SO2JalcamNoch keine Bewertungen
- Método para Determinar AmilosaDokument5 SeitenMétodo para Determinar AmilosaJalcamNoch keine Bewertungen
- Drones y Celdas de HidrogenoDokument11 SeitenDrones y Celdas de HidrogenoJalcamNoch keine Bewertungen
- Valorizacion Del Desecho de Aguas Residuales Del Prosamiento de AceitunasDokument11 SeitenValorizacion Del Desecho de Aguas Residuales Del Prosamiento de AceitunasJalcamNoch keine Bewertungen
- Diseño de Una Celda Microbiana de Combustible y Su Transición A Celda Microbiana de Electrólisis para Producción de Hidrógeno Por ElectrohidrogenesisDokument6 SeitenDiseño de Una Celda Microbiana de Combustible y Su Transición A Celda Microbiana de Electrólisis para Producción de Hidrógeno Por ElectrohidrogenesisJalcamNoch keine Bewertungen
- Composición Química de Aceitunas Durante La FermentacionDokument8 SeitenComposición Química de Aceitunas Durante La FermentacionJalcamNoch keine Bewertungen
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Role of Metallurgy in Mechanical EngineeringDokument4 SeitenRole of Metallurgy in Mechanical EngineeringMUHAMAD HAMMAD AHMEDNoch keine Bewertungen
- Ferroxyl Test Set: Vecom MarineDokument2 SeitenFerroxyl Test Set: Vecom MarineMarcelino Navarrete EvaristoNoch keine Bewertungen
- Lipofer Technical Report by LipofoodsDokument16 SeitenLipofer Technical Report by LipofoodsFernando HuaytaNoch keine Bewertungen
- Tribo Behav PTFE Sliding Against Steel Sea Water PDFDokument8 SeitenTribo Behav PTFE Sliding Against Steel Sea Water PDFarmin_kunosic9871Noch keine Bewertungen
- ReferencesDokument25 SeitenReferencesChriaAkaleziNoch keine Bewertungen
- The Mechanisms of Formation of W e L D Defects in High-Frequency Electric Resistance W e L D I N GDokument6 SeitenThe Mechanisms of Formation of W e L D Defects in High-Frequency Electric Resistance W e L D I N Gilze86Noch keine Bewertungen
- Clinker ReductionDokument2 SeitenClinker ReductionGiequatNoch keine Bewertungen
- Iron-Carbon Phase Diagram (SubsTech)Dokument2 SeitenIron-Carbon Phase Diagram (SubsTech)Aboo BackerNoch keine Bewertungen
- 2019 Hamdan Et Al Int J PhytoremediationDokument9 Seiten2019 Hamdan Et Al Int J Phytoremediationfiradila ainunnisaNoch keine Bewertungen
- Fe SADokument5 SeitenFe SASashi ThilakNoch keine Bewertungen
- Iron Carbon Phase DiagramDokument3 SeitenIron Carbon Phase DiagramrabikmNoch keine Bewertungen
- ReportDokument28 SeitenReportDipanshu KumarNoch keine Bewertungen
- Hom Sc. Apti PlusDokument560 SeitenHom Sc. Apti PlusSidharth BhoiNoch keine Bewertungen
- E-GMAT SC Hard Answer ExpDokument76 SeitenE-GMAT SC Hard Answer Expsumana i100% (1)
- Metals and Non Metals QuestionsDokument2 SeitenMetals and Non Metals QuestionsraghavNoch keine Bewertungen
- Response of Key Soil Parameters During Compost-Assisted Phytostabilization in Extremely Acidic Tailings: Effect of Plant SpeciesDokument9 SeitenResponse of Key Soil Parameters During Compost-Assisted Phytostabilization in Extremely Acidic Tailings: Effect of Plant Speciesgor45Noch keine Bewertungen
- The Iron-Iron Carbide (Fe-Fe3C) PhaseDokument8 SeitenThe Iron-Iron Carbide (Fe-Fe3C) PhaseRecep VatanseverNoch keine Bewertungen
- Balancing Chemical Equations Worksheet Student Instructions: Acids Alkalis (Bases)Dokument5 SeitenBalancing Chemical Equations Worksheet Student Instructions: Acids Alkalis (Bases)Gopal PenjarlaNoch keine Bewertungen
- Tribological and Mechanical Characterization of Nickel Aluminium Bronze (NAB) Manufactured by Laser Powder-Bed Fusion (L-PBF)Dokument10 SeitenTribological and Mechanical Characterization of Nickel Aluminium Bronze (NAB) Manufactured by Laser Powder-Bed Fusion (L-PBF)Fathia AlkelaeNoch keine Bewertungen
- Ferric Sulphate 112 Specification Rev 4Dokument1 SeiteFerric Sulphate 112 Specification Rev 4Edith WangNoch keine Bewertungen
- Worldwide Steel GradeDokument2 SeitenWorldwide Steel Grademyusuf123Noch keine Bewertungen
- Soal KimiaDokument8 SeitenSoal KimiaNindya Galuh KartikawatiNoch keine Bewertungen
- 2brick Properties and ManufacturingDokument37 Seiten2brick Properties and ManufacturingGaurav KandelNoch keine Bewertungen
- Chapter 1 Chemical Reactions and EquationsDokument12 SeitenChapter 1 Chemical Reactions and EquationsPrabhuPalanichamyNoch keine Bewertungen
- Metals, Non Metals, and MetallurgyDokument13 SeitenMetals, Non Metals, and MetallurgyM. Amebari NongsiejNoch keine Bewertungen
- Soalan Kimia Pertengahan Tahun Form 4Dokument11 SeitenSoalan Kimia Pertengahan Tahun Form 4Ridzuan Mohd AliNoch keine Bewertungen
- Nickel Troubleshooting GuideDokument45 SeitenNickel Troubleshooting GuideFrancisco BocanegraNoch keine Bewertungen
- 153-Paleoenvironmental Interpretation of Last Millennium Sediments in The Marriaga Swamp, Atrato Delta, Colombia.-2021Dokument16 Seiten153-Paleoenvironmental Interpretation of Last Millennium Sediments in The Marriaga Swamp, Atrato Delta, Colombia.-2021vere blancoNoch keine Bewertungen
- Mram 2 Dieny PDFDokument25 SeitenMram 2 Dieny PDFGokaran ShuklaNoch keine Bewertungen
- 14250A ch1 PDFDokument25 Seiten14250A ch1 PDFnagarajNoch keine Bewertungen