Das könnte Ihnen auch gefallen
- Artigo-Aditivos Alimentares-Aminoacidos e Vitaminas PDFDokument11 SeitenArtigo-Aditivos Alimentares-Aminoacidos e Vitaminas PDFPriscilla SuassunaNoch keine Bewertungen
- CarotenoidesDokument16 SeitenCarotenoidesLuanaSiqueira100% (1)
- Produção de Acidos OrganicosDokument6 SeitenProdução de Acidos OrganicosdcvidaNoch keine Bewertungen
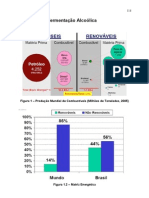
- 7 - Fermentação AlcoólicaDokument85 Seiten7 - Fermentação AlcoólicalabaptisNoch keine Bewertungen
- Biotecnologia Aplicada A Alimentação de Não Ruminantes - WilsonDokument18 SeitenBiotecnologia Aplicada A Alimentação de Não Ruminantes - WilsonWilson SilvaNoch keine Bewertungen
- Produção Industrial de Ácido Citrico Por Yarrowia LipolylicaDokument21 SeitenProdução Industrial de Ácido Citrico Por Yarrowia LipolylicaMariana MichelNoch keine Bewertungen
- Aps - Aplicação Da Cinética Enzimática Nos Processos Farmacêuticos FermentativosDokument14 SeitenAps - Aplicação Da Cinética Enzimática Nos Processos Farmacêuticos FermentativosLucy ModoloNoch keine Bewertungen
- Cultivo de Cogumelos em Residuos de CervejaDokument25 SeitenCultivo de Cogumelos em Residuos de CervejaBruno GadêlhaNoch keine Bewertungen
- 7653 Microbiologia Industrial e BiotecnologiaDokument17 Seiten7653 Microbiologia Industrial e BiotecnologiaCarlos Henrique Machado DiasNoch keine Bewertungen
- Ana769lise Patentome769trica e Bibliome769trica 2020Dokument8 SeitenAna769lise Patentome769trica e Bibliome769trica 2020Narraque GarciaNoch keine Bewertungen
- Produção de Medicamentos Com o Uso de MicrorganismosDokument13 SeitenProdução de Medicamentos Com o Uso de MicrorganismosBruno Carvalho SilvaNoch keine Bewertungen
- 05 - Biotecnologia Aplicada Ao Valor Nutricional Dos AlimentosDokument8 Seiten05 - Biotecnologia Aplicada Ao Valor Nutricional Dos AlimentosFernanda OliveiraNoch keine Bewertungen
- Aminoacidos Nas Plantas 20210510161940Dokument7 SeitenAminoacidos Nas Plantas 20210510161940cleidianeNoch keine Bewertungen
- Aula - Metabolismo de CarboidratosDokument24 SeitenAula - Metabolismo de CarboidratosRicardo GomesNoch keine Bewertungen
- Crescimento e Metabolismo MicrobianoDokument87 SeitenCrescimento e Metabolismo Microbianovitor battistella mayNoch keine Bewertungen
- Artigo Julho 2021 3Dokument14 SeitenArtigo Julho 2021 3VanMarvasNoch keine Bewertungen
- Vitaminas LipossuluveisDokument15 SeitenVitaminas LipossuluveisNeli MatsinheNoch keine Bewertungen
- Biotecnologia IndustrialDokument10 SeitenBiotecnologia IndustrialEdgard De MoraesNoch keine Bewertungen
- Aula - 2 - Conceitos Importantes e AplicaçõesDokument12 SeitenAula - 2 - Conceitos Importantes e AplicaçõesAlecksia EvillynNoch keine Bewertungen
- Boletim 2-3Dokument52 SeitenBoletim 2-3Catarina SousaNoch keine Bewertungen
- Estudo Dirigido Enzimologia Segunda Etapa 2019 02Dokument2 SeitenEstudo Dirigido Enzimologia Segunda Etapa 2019 02Debora Costa ReisNoch keine Bewertungen
- Molecules 27 00410 V2.en - PTDokument16 SeitenMolecules 27 00410 V2.en - PTALEXANDRE SOUSANoch keine Bewertungen
- Soja, Aspectos Químicos E Classificação 1.1 ContextualizaçãoDokument6 SeitenSoja, Aspectos Químicos E Classificação 1.1 ContextualizaçãoWagner Gomes LopesNoch keine Bewertungen
- Bioquímica Ruminal PDFDokument21 SeitenBioquímica Ruminal PDFLuciana VianaNoch keine Bewertungen
- OTIMIZAÇÃO DO MEIO DE CULTIVO SUBMERSO DE Pseudomonas Aeruginosa UTILIZANDO RESIDUOS AGROINDUTRIAIS PARA PRODUÇÃO DE LIPASEDokument8 SeitenOTIMIZAÇÃO DO MEIO DE CULTIVO SUBMERSO DE Pseudomonas Aeruginosa UTILIZANDO RESIDUOS AGROINDUTRIAIS PARA PRODUÇÃO DE LIPASEJéssyca CiprianoNoch keine Bewertungen
- Artigo Sobre Subproduto Da SojaDokument7 SeitenArtigo Sobre Subproduto Da SojaBlenda SoaresNoch keine Bewertungen
- Cultivo Da Microalga Spirulina Platensis em FontesDokument7 SeitenCultivo Da Microalga Spirulina Platensis em FontesO ReikianoNoch keine Bewertungen
- Atividade Reflexiva - Feita LucianaDokument3 SeitenAtividade Reflexiva - Feita LucianalucianareginadeoliveiraoliveirNoch keine Bewertungen
- Ulva lactuca: Aspectos Químicos na Determinação da Atividade Antioxidante TotalVon EverandUlva lactuca: Aspectos Químicos na Determinação da Atividade Antioxidante TotalNoch keine Bewertungen
- Lista 2 - Bioquímica IndustrialDokument13 SeitenLista 2 - Bioquímica IndustrialGabriel CamposNoch keine Bewertungen
- Poligrafo Eng - 2016Dokument85 SeitenPoligrafo Eng - 2016Miriam CostaNoch keine Bewertungen
- Admin2,+01 444 2603 1 SPDokument8 SeitenAdmin2,+01 444 2603 1 SPANTONIO CELSO R DE OLIVEIRANoch keine Bewertungen
- Microbiologia IndustrialDokument6 SeitenMicrobiologia IndustrialMaria VitoriaNoch keine Bewertungen
- Produção de Plásticos Biodegradáveis Por Fermentação (PHA e PHB)Dokument27 SeitenProdução de Plásticos Biodegradáveis Por Fermentação (PHA e PHB)Lorraine OliveiraNoch keine Bewertungen
- Processos FermentativosDokument50 SeitenProcessos FermentativosStefanya De Paula FreitasNoch keine Bewertungen
- Microalgas Na Produção de Biodiesel: Extração e Composição LipídicaDokument8 SeitenMicroalgas Na Produção de Biodiesel: Extração e Composição LipídicaJonathas FerreiraNoch keine Bewertungen
- VOL3-CAP 19 Fermentação Alcoólica RaízesDokument23 SeitenVOL3-CAP 19 Fermentação Alcoólica RaízesLamar MorenoNoch keine Bewertungen
- Biossíntese - TrabalhoDokument46 SeitenBiossíntese - TrabalhogaperimNoch keine Bewertungen
- Artigo Qualidade Do OvoDokument10 SeitenArtigo Qualidade Do OvoCarina TrevisanNoch keine Bewertungen
- Cogumelos: Uma Fonte Promissora de Compostos Ativos para O Desenvolvimento de Bioprodutos Farmacêuticos E NutracêuticosDokument46 SeitenCogumelos: Uma Fonte Promissora de Compostos Ativos para O Desenvolvimento de Bioprodutos Farmacêuticos E NutracêuticosAnonymous Kti5jq5EJINoch keine Bewertungen
- Microorganismos de Uso BiotecnológicosDokument3 SeitenMicroorganismos de Uso BiotecnológicosAmanda LanzottiNoch keine Bewertungen
- Artigo Produção de Biogás A Partir de Dejetos de SuínosDokument12 SeitenArtigo Produção de Biogás A Partir de Dejetos de SuínosMauro OrcelliNoch keine Bewertungen
- Biotec e MicrobiologiaDokument23 SeitenBiotec e MicrobiologiaTHEUS SNOW'SNoch keine Bewertungen
- Manual Biodigestor WinrockDokument21 SeitenManual Biodigestor WinrockrlsaitoNoch keine Bewertungen
- Relatório - Purificação de Proteina - Produção e Purificação de LipaseDokument23 SeitenRelatório - Purificação de Proteina - Produção e Purificação de LipaseDanilo José100% (1)
- Apostila - Aula 1Dokument21 SeitenApostila - Aula 1Vitor MonteiroNoch keine Bewertungen
- Produção de BioinseticidasDokument26 SeitenProdução de BioinseticidasMayara Castro de MoraisNoch keine Bewertungen
- Etanol de biomassa de milho: utilização de aprendizagem de máquina no estudo de casos de caldeirasVon EverandEtanol de biomassa de milho: utilização de aprendizagem de máquina no estudo de casos de caldeirasNoch keine Bewertungen
- Processo Fermentativo, Digestivo e Fatores Antinutricionais de Nutrientes para RuminantesDokument13 SeitenProcesso Fermentativo, Digestivo e Fatores Antinutricionais de Nutrientes para RuminantesLuiz FelipeNoch keine Bewertungen
- Aula 1 - Processos Fermentativo IndustrialDokument30 SeitenAula 1 - Processos Fermentativo Industrialxinha005Noch keine Bewertungen
- Bioethanol Production Using Microalgae A ReviewDokument16 SeitenBioethanol Production Using Microalgae A ReviewLAURA LUC�A ATENCIA CASTILLONoch keine Bewertungen
- Fermentação Contínua e DescontinuaDokument11 SeitenFermentação Contínua e DescontinuaLi Shauon0% (1)
- BiogasDokument2 SeitenBiogaspaulo monteiroNoch keine Bewertungen
- Admin2, 104Dokument24 SeitenAdmin2, 104Teresa MendoncaNoch keine Bewertungen
- Estado Da Arte Da Produção de Biogás A Partir Dos Resíduos Do Processamento Da MandiocaDokument21 SeitenEstado Da Arte Da Produção de Biogás A Partir Dos Resíduos Do Processamento Da MandiocaAdriano Henrique FerrarezNoch keine Bewertungen
- Operações Unitárias na Indústria de Alimentos e QuímicaVon EverandOperações Unitárias na Indústria de Alimentos e QuímicaNoch keine Bewertungen
- A Utilização De Material Lignocelulósico Na Produção De BioetanolVon EverandA Utilização De Material Lignocelulósico Na Produção De BioetanolNoch keine Bewertungen
- Relatório Leito Fixo e FluidizadoDokument29 SeitenRelatório Leito Fixo e FluidizadoElias JúniorNoch keine Bewertungen
- Indústria Alcoolquímica - Elias e Thaís PDFDokument12 SeitenIndústria Alcoolquímica - Elias e Thaís PDFElias JúniorNoch keine Bewertungen
- Lista06 - Mecânica Das Estruturas IDokument7 SeitenLista06 - Mecânica Das Estruturas IElias JúniorNoch keine Bewertungen
- Relatório LEQI - Leito Fixo e FluidizadoDokument30 SeitenRelatório LEQI - Leito Fixo e FluidizadoElias JúniorNoch keine Bewertungen
- RDC 51 e 52Dokument5 SeitenRDC 51 e 52Elias JúniorNoch keine Bewertungen
- JoseJorgeMarques - O Plástico Verde e o Mercado Brasileiro de Etanol PDFDokument154 SeitenJoseJorgeMarques - O Plástico Verde e o Mercado Brasileiro de Etanol PDFElias JúniorNoch keine Bewertungen
- Relatorio - Trocador de CalorDokument24 SeitenRelatorio - Trocador de CalorElias JúniorNoch keine Bewertungen
- NBR 06400 MB 1032 - Bombas Hidraulicas de Fluxo (Classe C) - Ensaios de Desempenho e de CavitacaoDokument26 SeitenNBR 06400 MB 1032 - Bombas Hidraulicas de Fluxo (Classe C) - Ensaios de Desempenho e de CavitacaoRenatoNoch keine Bewertungen
- Relatorio 3 - Trocador de Calor - CORRIGIDO PDFDokument24 SeitenRelatorio 3 - Trocador de Calor - CORRIGIDO PDFElias JúniorNoch keine Bewertungen
- InsetosDokument5 SeitenInsetosElias JúniorNoch keine Bewertungen
- GUIA de Fórmulas SkinCare para Salvar No Celular e Computador PDFDokument137 SeitenGUIA de Fórmulas SkinCare para Salvar No Celular e Computador PDFrenata andrade100% (5)
- 1 Pedologia LivroDokument94 Seiten1 Pedologia LivroFilipe MacielNoch keine Bewertungen
- Obtenção Do OxigênioDokument4 SeitenObtenção Do OxigênioAndre MatsumotoNoch keine Bewertungen
- O Estresse Causado Pela Aceleração Dos PensamentosDokument3 SeitenO Estresse Causado Pela Aceleração Dos PensamentosCelia GevartoskiNoch keine Bewertungen
- Disbiose Intestinal Canina Diagnostico e TratamentoDokument8 SeitenDisbiose Intestinal Canina Diagnostico e Tratamentohel-lopesNoch keine Bewertungen
- Autismo - PDF Intervencao-Nutricional-No - (4699)Dokument36 SeitenAutismo - PDF Intervencao-Nutricional-No - (4699)Taiana Cardoso100% (1)
- Aula Anatomia VegetalDokument8 SeitenAula Anatomia VegetalLEMOSNoch keine Bewertungen
- Projeto de Laboratório BiomédicoDokument16 SeitenProjeto de Laboratório BiomédicoCarlos Guilherme MaquinéNoch keine Bewertungen
- Tecnica Alternativa para Montagem de Insetos em LaminasDokument7 SeitenTecnica Alternativa para Montagem de Insetos em LaminasfebarlettaNoch keine Bewertungen
- Relacionamentos Amorosos Na JuventudeDokument20 SeitenRelacionamentos Amorosos Na JuventudeEliezer BordinhãoNoch keine Bewertungen
- Agrometeorologia EvapotranspiraçãoDokument17 SeitenAgrometeorologia EvapotranspiraçãoDAIENE FRANCIELLI SANTOSNoch keine Bewertungen
- DorsalDokument3 SeitenDorsalFabianinhoLacerdaNoch keine Bewertungen
- Inteligencia EmocionalDokument460 SeitenInteligencia EmocionalMárcia MartinsNoch keine Bewertungen
- O Cérebro DesconhecidoDokument7 SeitenO Cérebro DesconhecidoBautista O Diana75% (4)
- 09 - Filhos, Provisão de DeusDokument0 Seiten09 - Filhos, Provisão de DeusAdv Val MmnNoch keine Bewertungen
- LICOPENODokument4 SeitenLICOPENOFrancine SallaNoch keine Bewertungen
- 4ListadoEnergia Gibbs HelmholtzDokument2 Seiten4ListadoEnergia Gibbs Helmholtzgustav23450% (2)
- 15 Dicas Infalíveis para Desanuviar A Sua Mente - Estado ZenDokument3 Seiten15 Dicas Infalíveis para Desanuviar A Sua Mente - Estado ZenArnaldo GomesNoch keine Bewertungen
- Noçoes de Biologia para PeríciaDokument48 SeitenNoçoes de Biologia para PeríciaLuiz MiguelNoch keine Bewertungen
- Dercy Goncalves DesciclopediaDokument54 SeitenDercy Goncalves DesciclopediaheattedNoch keine Bewertungen
- 7 Grupo QF 1Dokument56 Seiten7 Grupo QF 1Gildo Salai MacalaneNoch keine Bewertungen
- Nos Dominios Da Mediunidade - Chico XavierDokument151 SeitenNos Dominios Da Mediunidade - Chico XavierMauricio Gonçalves100% (1)
- Princípios Da VirologiaDokument49 SeitenPrincípios Da VirologiaAna Paula Do Nascimento100% (1)
- Slide NR 6Dokument35 SeitenSlide NR 6Luciano Joviano0% (1)
- Jaymin Eve - SuperNatural Prison Story 01 - Broken CompassDokument295 SeitenJaymin Eve - SuperNatural Prison Story 01 - Broken CompassRobson Magno100% (1)
- Artigo - O Potencial de Membrana - Khan AcademyDokument26 SeitenArtigo - O Potencial de Membrana - Khan AcademyJéssica LeãoNoch keine Bewertungen
- 2018 SANTAELLAArte Cienciae Tecnologia Campo ExpansaoDokument24 Seiten2018 SANTAELLAArte Cienciae Tecnologia Campo ExpansaoLoren BergantiniNoch keine Bewertungen
- Distúrbios Da Consciência Humana - Parte 2 de 3 A Abordagem Dos Enfermos em ComaDokument8 SeitenDistúrbios Da Consciência Humana - Parte 2 de 3 A Abordagem Dos Enfermos em ComaAyrk ZamiskeNoch keine Bewertungen
- A Percepção de Riscos e A Redução Dos Acidentes de TrabalhoDokument34 SeitenA Percepção de Riscos e A Redução Dos Acidentes de TrabalhoAntonio Fernando Navarro100% (1)
- 6oano-1b 2-Mat ALDokument55 Seiten6oano-1b 2-Mat ALEIDI BerataNoch keine Bewertungen