Das könnte Ihnen auch gefallen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Nesters Microbiology A Human Perspective 9th Edition Anderson Test BankDokument25 SeitenNesters Microbiology A Human Perspective 9th Edition Anderson Test BankRicardoPetersJrdstf100% (51)
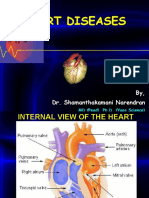
- Cardio Vascular DiseaseDokument86 SeitenCardio Vascular Diseasemanju09535Noch keine Bewertungen
- TradingDokument5 SeitenTradingmanju09535Noch keine Bewertungen
- Why Is Thymine Replaced by Uracil in RNADokument2 SeitenWhy Is Thymine Replaced by Uracil in RNAmanju09535Noch keine Bewertungen
- HR Project.Dokument76 SeitenHR Project.manju09535Noch keine Bewertungen
- Annexure Ii Affidavit by Parent/GuardianDokument2 SeitenAnnexure Ii Affidavit by Parent/Guardianmanju09535Noch keine Bewertungen
- Technical AnalysisDokument16 SeitenTechnical Analysismanju09535Noch keine Bewertungen
- 24 12 June 2012Dokument2 Seiten24 12 June 2012manju09535Noch keine Bewertungen
- Income Under The Head Capital Gains' and Its ComputationDokument12 SeitenIncome Under The Head Capital Gains' and Its Computationmanju09535Noch keine Bewertungen
- English GrammerDokument29 SeitenEnglish Grammermanju09535Noch keine Bewertungen
- ExeclDokument28 SeitenExeclmanju09535Noch keine Bewertungen
- Excel Comments - Basics: Insert An Excel CommentDokument13 SeitenExcel Comments - Basics: Insert An Excel Commentmanju09535Noch keine Bewertungen
- Project Appraisal - Capital Budgeting MethodsDokument55 SeitenProject Appraisal - Capital Budgeting Methodsmanju09535Noch keine Bewertungen
- Electrical Properties of Materials: Chap 1Dokument28 SeitenElectrical Properties of Materials: Chap 1Shuvro Sankar SenNoch keine Bewertungen
- Lecture 04 - Bipolar Junction Transistors (BJT)Dokument39 SeitenLecture 04 - Bipolar Junction Transistors (BJT)Joseph NgowiNoch keine Bewertungen
- Photoelectric EffectDokument4 SeitenPhotoelectric EffectMatheus Loss LizeNoch keine Bewertungen
- Mettalic BondDokument32 SeitenMettalic BondKim Paolo Armin Torcelino VINoch keine Bewertungen
- Syllabus Quantum MechanicsDokument7 SeitenSyllabus Quantum MechanicsRidika ChaturvediNoch keine Bewertungen
- Crystal Field Theory - NURDokument5 SeitenCrystal Field Theory - NURNurhajrahNoch keine Bewertungen
- NET QuestionsDokument3 SeitenNET QuestionsSouvik DasNoch keine Bewertungen
- Eutectic Mixtures: Similarities Between Eutectic Mixture and Pure CompoundDokument8 SeitenEutectic Mixtures: Similarities Between Eutectic Mixture and Pure CompoundNaduku EridadNoch keine Bewertungen
- Structure of The f6 Configuration With Application To Rare Earth IonsDokument11 SeitenStructure of The f6 Configuration With Application To Rare Earth Ionsw_i_tNoch keine Bewertungen
- Magnetic PropertiesDokument3 SeitenMagnetic PropertiesAly HannahNoch keine Bewertungen
- Nano Chemistry 2Dokument11 SeitenNano Chemistry 2N AkilanNoch keine Bewertungen
- Lecture 2-Crystal StructureDokument27 SeitenLecture 2-Crystal StructureAtika AlamNoch keine Bewertungen
- SIMS Analysis of A Multiple Quantum Well Structure in A Vertical Cavity Surface Emitting Laser Using The Mixing-Roughness-Information Depth ModelDokument4 SeitenSIMS Analysis of A Multiple Quantum Well Structure in A Vertical Cavity Surface Emitting Laser Using The Mixing-Roughness-Information Depth Modelsepot24093Noch keine Bewertungen
- Integrated Electronics - Millman HalkiasDokument957 SeitenIntegrated Electronics - Millman HalkiasLEKSHMI ANILKUMARNoch keine Bewertungen
- Spin Valve TransistorDokument22 SeitenSpin Valve Transistorkandimalla_rajaneeshNoch keine Bewertungen
- Master of Science (M.SC.) (Physics) Semester-IV (C.B.S.) Examination Solid State Physics-Ii Compulsory Paper-1Dokument2 SeitenMaster of Science (M.SC.) (Physics) Semester-IV (C.B.S.) Examination Solid State Physics-Ii Compulsory Paper-1Sanyam KumariNoch keine Bewertungen
- Chapter Six: X-RaysDokument16 SeitenChapter Six: X-RaysAhmed ZagharyNoch keine Bewertungen
- Perovskyt 5 ADokument18 SeitenPerovskyt 5 ASegundo AcevedoNoch keine Bewertungen
- Chemical BondingDokument38 SeitenChemical Bondingsmileworker utkarshsinghNoch keine Bewertungen
- Semiconductor Interview Questions and Answers 2022Dokument6 SeitenSemiconductor Interview Questions and Answers 2022Naznin Nuria AfrinNoch keine Bewertungen
- (Physics of Solids and Liquids) N. H. March (Auth.), S. Lundqvist, N. H. March (Eds.) - Theory of The Inhomogeneous Electron Gas-Springer US (1983)Dokument405 Seiten(Physics of Solids and Liquids) N. H. March (Auth.), S. Lundqvist, N. H. March (Eds.) - Theory of The Inhomogeneous Electron Gas-Springer US (1983)Nicolas LizarraldeNoch keine Bewertungen
- DPP#21-22 PERIODIC TABLE Prev. YrDokument4 SeitenDPP#21-22 PERIODIC TABLE Prev. YrLakshya ChandakNoch keine Bewertungen
- Matlab/Pspice Hybrid Simulation Modeling of A Photovoltaic System and Its ApplicationsDokument137 SeitenMatlab/Pspice Hybrid Simulation Modeling of A Photovoltaic System and Its ApplicationsJaafar AbbakarNoch keine Bewertungen
- Investigation On The Crystallinity and Optoelectronic Properties of Thin Films of Zno/mapbx3heterojunctionsynthesized Via The Spin-Coating MethodDokument8 SeitenInvestigation On The Crystallinity and Optoelectronic Properties of Thin Films of Zno/mapbx3heterojunctionsynthesized Via The Spin-Coating MethodIJAR JOURNALNoch keine Bewertungen
- Quantum Dot Laser Full ReportDokument38 SeitenQuantum Dot Laser Full ReportRajeev Sndhr50% (2)
- 3-D Solar Cells by Electrochemical-Deposited Se LaDokument8 Seiten3-D Solar Cells by Electrochemical-Deposited Se LaIndrajit ShriwastavNoch keine Bewertungen
- Chem350 Notes-6-20111Dokument13 SeitenChem350 Notes-6-20111Sara FatimaNoch keine Bewertungen
- Dipoles and Intermolecular ForcesDokument14 SeitenDipoles and Intermolecular Forcesspidy warriorsNoch keine Bewertungen
- QUARTER 2 Performance TaskDokument8 SeitenQUARTER 2 Performance TaskJaime CrispinoNoch keine Bewertungen