Das könnte Ihnen auch gefallen
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Wilderness Trails Zoo Coloring Activity BookDokument24 SeitenWilderness Trails Zoo Coloring Activity BookHolly Crews100% (2)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Isotonic, Hypotonic, Hypertonic IV Third Spacing Where Has All The Fluid GoneDokument14 SeitenIsotonic, Hypotonic, Hypertonic IV Third Spacing Where Has All The Fluid GoneJMC5053100% (2)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- Methionine PDFDokument20 SeitenMethionine PDFChamara SamaraweeraNoch keine Bewertungen
- Arabic Hebrew Greek Study SheetDokument75 SeitenArabic Hebrew Greek Study SheetAnonymous 7QjNuvoCpINoch keine Bewertungen
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Judy Bolton #31 The Discovery at The Dragon's MouthDokument200 SeitenJudy Bolton #31 The Discovery at The Dragon's MouthPastPresentFuture100% (1)
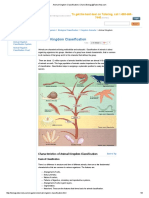
- Animal Kingdom Classification - Chart - Biology@TutorVistaDokument6 SeitenAnimal Kingdom Classification - Chart - Biology@TutorVistaAnujith AnuNoch keine Bewertungen
- Basic Life SupportDokument9 SeitenBasic Life Supportaina_boydonNoch keine Bewertungen
- Microblading ManualDokument6 SeitenMicroblading ManualPaulina Pineda100% (1)
- Digestive System of The FrogDokument73 SeitenDigestive System of The FrogNi Ki TaNoch keine Bewertungen
- MENOPAUSEDokument27 SeitenMENOPAUSEKarishma Shroff83% (6)
- Irregular Plurals ListDokument7 SeitenIrregular Plurals ListAnonymous OQmcDMgNoch keine Bewertungen
- Obesity - The Built EnvironmentDokument2 SeitenObesity - The Built EnvironmentvcoglianeseNoch keine Bewertungen
- Obesity - Stigma of ObesityDokument2 SeitenObesity - Stigma of ObesityvcoglianeseNoch keine Bewertungen
- Obesity - Hunter GathererDokument1 SeiteObesity - Hunter GatherervcoglianeseNoch keine Bewertungen
- Endocannabinoid Control of Food Intake and Energy Balance (Marzo & Matias)Dokument2 SeitenEndocannabinoid Control of Food Intake and Energy Balance (Marzo & Matias)vcoglianeseNoch keine Bewertungen
- Obesity - Decreased Fatty Acid OxidationDokument2 SeitenObesity - Decreased Fatty Acid OxidationvcoglianeseNoch keine Bewertungen
- Loaf Cake or Cup Cakes: Pumpkin Spice Pudding Bundt CakeDokument3 SeitenLoaf Cake or Cup Cakes: Pumpkin Spice Pudding Bundt CakevcoglianeseNoch keine Bewertungen
- Dietary Guidelines: ExecSummDokument4 SeitenDietary Guidelines: ExecSummPatricia DillonNoch keine Bewertungen
- Structure Revisi Baru - TrawasDokument76 SeitenStructure Revisi Baru - TrawasEdy KurniawanNoch keine Bewertungen
- Excellup Class TenDokument3 SeitenExcellup Class TenHemant KumarNoch keine Bewertungen
- Anaphy Topic 3 IntegralDokument3 SeitenAnaphy Topic 3 Integral박제라Noch keine Bewertungen
- GHOP Menu 2012 2 24Dokument2 SeitenGHOP Menu 2012 2 24administrator8147Noch keine Bewertungen
- Answers WSKHT Lethal DoseDokument2 SeitenAnswers WSKHT Lethal DosejdNoch keine Bewertungen
- Adaptive Radiation and Speciation LabDokument2 SeitenAdaptive Radiation and Speciation LabzacharyNoch keine Bewertungen
- Changing Places Dominoes 1Dokument2 SeitenChanging Places Dominoes 1Lucila Fernández SalcedoNoch keine Bewertungen
- Timms 1980Dokument8 SeitenTimms 1980Zahid ManzoorNoch keine Bewertungen
- NEC 11th Five Year Plan 2007-2011Dokument109 SeitenNEC 11th Five Year Plan 2007-2011Kamzalian Tomging100% (3)
- Simple Garter Stitch Slippers v2Dokument3 SeitenSimple Garter Stitch Slippers v2sylviacristinariveraNoch keine Bewertungen
- Eng-B w10 InternetDokument6 SeitenEng-B w10 InternetmoralassNoch keine Bewertungen
- 3-John Gerard Possible Defects in Leather Production PDFDokument379 Seiten3-John Gerard Possible Defects in Leather Production PDFAwanis LuthfanNoch keine Bewertungen
- Integumentary Lab ActDokument5 SeitenIntegumentary Lab ActJASMINE LAURONNoch keine Bewertungen
- 2023 CBSE I Succeed Eng. Lang. Lit. K. A. Zone Poetry SectionDokument9 Seiten2023 CBSE I Succeed Eng. Lang. Lit. K. A. Zone Poetry SectioniCOS iSpace100% (1)
- Brown Garden SnailDokument6 SeitenBrown Garden SnailyaaqootNoch keine Bewertungen
- Phi002 Lesson 9Dokument5 SeitenPhi002 Lesson 9Monique EamiguelNoch keine Bewertungen
- Life Cycle of A FrogDokument2 SeitenLife Cycle of A FrogMary Grace YañezNoch keine Bewertungen
- Turn033016web PDFDokument28 SeitenTurn033016web PDFtodaysshopperNoch keine Bewertungen
- Second Cot Science SixDokument5 SeitenSecond Cot Science Sixmaricel isletaNoch keine Bewertungen