Das könnte Ihnen auch gefallen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Statistical Tolerance Stack AnalysisDokument52 SeitenStatistical Tolerance Stack Analysisdineshnexus100% (1)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- Hawking-Brief History of TimeDokument336 SeitenHawking-Brief History of TimeAlbert Kristian0% (1)
- Ansys Theory ReferenceDokument1.067 SeitenAnsys Theory ReferencepilafaNoch keine Bewertungen
- Delphi 9322a000Dokument5 SeitenDelphi 9322a000BaytolgaNoch keine Bewertungen
- Sip TrainingDokument96 SeitenSip Trainingronics123Noch keine Bewertungen
- Java10 PDFDokument137 SeitenJava10 PDFswarup sarkarNoch keine Bewertungen
- Cds 13 WorkbookDokument99 SeitenCds 13 WorkbookWaqar A. KhanNoch keine Bewertungen
- Lafortune - Et - Al - SMST06 - Superelasticity Related Phenomena of Shape Memory Alloy Passive DampersDokument10 SeitenLafortune - Et - Al - SMST06 - Superelasticity Related Phenomena of Shape Memory Alloy Passive DamperspilafaNoch keine Bewertungen
- Elmer Finite Element Software for Multiphysical Optimization ProblemsDokument28 SeitenElmer Finite Element Software for Multiphysical Optimization ProblemssyammohansNoch keine Bewertungen
- High Performance Computational Electromechanical Model of The HeartDokument4 SeitenHigh Performance Computational Electromechanical Model of The HeartpilafaNoch keine Bewertungen
- A Comparative Thermal Study of Two Motors StructureDokument15 SeitenA Comparative Thermal Study of Two Motors StructurepilafaNoch keine Bewertungen
- GT&D GlossaryDokument10 SeitenGT&D GlossaryshawntsungNoch keine Bewertungen
- Easily Made Errors in FEA 1Dokument6 SeitenEasily Made Errors in FEA 1darkwing888Noch keine Bewertungen
- Dynamic Non-Linear Electro-Thermal Simulation of A Thin-Film Thermal ConverterDokument8 SeitenDynamic Non-Linear Electro-Thermal Simulation of A Thin-Film Thermal ConverterpilafaNoch keine Bewertungen
- FEMAP 2193 - tcm903-5012Dokument16 SeitenFEMAP 2193 - tcm903-5012Roger JiménezNoch keine Bewertungen
- Salome: The Open Source Integration Platform For Numerical SimulationDokument8 SeitenSalome: The Open Source Integration Platform For Numerical SimulationpilafaNoch keine Bewertungen
- Ansys Solver 2002Dokument62 SeitenAnsys Solver 2002pilafaNoch keine Bewertungen
- Elmer Solver ManualDokument131 SeitenElmer Solver ManualpilafaNoch keine Bewertungen
- Elmer TutorialsDokument103 SeitenElmer TutorialsChandrajit ThaokarNoch keine Bewertungen
- Parallel Computing With ElmerDokument54 SeitenParallel Computing With ElmerpilafaNoch keine Bewertungen
- Modal AnalysisDokument14 SeitenModal Analysismichael_r_reid652Noch keine Bewertungen
- Mat C ManualDokument8 SeitenMat C ManualBi8ikityNoch keine Bewertungen
- Large Scale Continuum Electromechanical Simulation of The HeartDokument1 SeiteLarge Scale Continuum Electromechanical Simulation of The HeartpilafaNoch keine Bewertungen
- High Performance Computational Electromechanical Model of The HeartDokument4 SeitenHigh Performance Computational Electromechanical Model of The HeartpilafaNoch keine Bewertungen
- Coupled Electromechanical Model of The Heart: Parallel Finite Element FormulationDokument15 SeitenCoupled Electromechanical Model of The Heart: Parallel Finite Element FormulationpilafaNoch keine Bewertungen
- An Introduction To Code AsterDokument71 SeitenAn Introduction To Code AsterIommoi MoiiomNoch keine Bewertungen
- 8086 Instruction SetDokument66 Seiten8086 Instruction SetRaj KumarNoch keine Bewertungen
- Velocity profiles and incompressible flow field equationsDokument2 SeitenVelocity profiles and incompressible flow field equationsAbdul ArifNoch keine Bewertungen
- Tips and Tricks I: Getting the Most Out of ArcGIS DesktopDokument108 SeitenTips and Tricks I: Getting the Most Out of ArcGIS Desktoptanja222Noch keine Bewertungen
- Ain 2016 Pilot Report m600Dokument4 SeitenAin 2016 Pilot Report m600Antonio Cesar de Sa LeitaoNoch keine Bewertungen
- Notifier Battery Calculations-ReadmeDokument11 SeitenNotifier Battery Calculations-ReadmeJeanCarlosRiveroNoch keine Bewertungen
- CS 102 Programming Fundamentals Lecture NotesDokument14 SeitenCS 102 Programming Fundamentals Lecture NotesOkay OkayNoch keine Bewertungen
- Unit 10Dokument18 SeitenUnit 10ChaithraMalluNoch keine Bewertungen
- Numeri OrdinaliDokument2 SeitenNumeri OrdinaliClaudioNoch keine Bewertungen
- GAS-INSULATED SWITCHGEAR MODELS 72kV ADVANCED ENVIRONMENTALLY FRIENDLYDokument6 SeitenGAS-INSULATED SWITCHGEAR MODELS 72kV ADVANCED ENVIRONMENTALLY FRIENDLYBudi SantonyNoch keine Bewertungen
- General 04 Fixed Flow Pump To Three TanksDokument13 SeitenGeneral 04 Fixed Flow Pump To Three TanksjpalauguillemNoch keine Bewertungen
- Midterm Exam Result Ce199-1l 2Q1920Dokument3 SeitenMidterm Exam Result Ce199-1l 2Q1920RA CarpioNoch keine Bewertungen
- 1/2" Cellflex Superflexible Foam-Dielectric Coaxial Cable: SCF12-50JDokument2 Seiten1/2" Cellflex Superflexible Foam-Dielectric Coaxial Cable: SCF12-50JpeguigonsoNoch keine Bewertungen
- Superalloy Brochure PDFDokument16 SeitenSuperalloy Brochure PDFDaren NeradNoch keine Bewertungen
- Logic CHPT71Dokument27 SeitenLogic CHPT71Eronjosh FontanozaNoch keine Bewertungen
- Answer Key Grade 6 - 2023Dokument7 SeitenAnswer Key Grade 6 - 2023ALKHANSAA ELOBEIDYNoch keine Bewertungen
- Service Manual: S&T Motors Co., LTDDokument94 SeitenService Manual: S&T Motors Co., LTDJuliano PedrosoNoch keine Bewertungen
- Front Panel & Display Technical Data: User ManualDokument2 SeitenFront Panel & Display Technical Data: User ManualJulio PorleyNoch keine Bewertungen
- 08 Candelaria Punta Del Cobre IOCG Deposits PDFDokument27 Seiten08 Candelaria Punta Del Cobre IOCG Deposits PDFDiego Morales DíazNoch keine Bewertungen
- QT140 500 KG Per Hr. Fish Feed Pelleting PlantDokument11 SeitenQT140 500 KG Per Hr. Fish Feed Pelleting PlantShekhar MitraNoch keine Bewertungen
- Rso PDFDokument120 SeitenRso PDFjohn shepardNoch keine Bewertungen
- EET422 EMC Intro-Banana Skins 2011-2012 MSWDokument6 SeitenEET422 EMC Intro-Banana Skins 2011-2012 MSWVeeradasan PerumalNoch keine Bewertungen
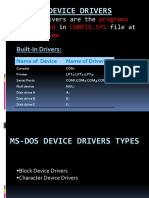
- Ms-Dos Device Drivers: Device Drivers Are The That in File atDokument13 SeitenMs-Dos Device Drivers: Device Drivers Are The That in File atJass GillNoch keine Bewertungen
- Camshaft Test SheetDokument4 SeitenCamshaft Test SheetsughieantoNoch keine Bewertungen
- Blowfish Encryption AlgorithmDokument3 SeitenBlowfish Encryption AlgorithmParkerAllisonNoch keine Bewertungen
- Scoop Atlas Wagner ST1810Dokument4 SeitenScoop Atlas Wagner ST1810Juan Manuel PerezNoch keine Bewertungen
- Introduction - Week 2Dokument37 SeitenIntroduction - Week 2Tayyab AhmedNoch keine Bewertungen