Das könnte Ihnen auch gefallen
- Guia de Desintoxicação de Proteína SpikeDokument21 SeitenGuia de Desintoxicação de Proteína SpikeGustavo VemnimimNoch keine Bewertungen
- FISPQ - Fumos Metalicos SoldaDokument23 SeitenFISPQ - Fumos Metalicos Soldaapi-3704990100% (4)
- 2020 - 04 GQ PT 2020 (Interactive) PDFDokument76 Seiten2020 - 04 GQ PT 2020 (Interactive) PDFTeresa PaivaNoch keine Bewertungen
- Cynara ScolymusDokument5 SeitenCynara Scolymusfernandojardim12Noch keine Bewertungen
- Cap 28 - Farmacologia Do TGIDokument6 SeitenCap 28 - Farmacologia Do TGIfernandojardim12Noch keine Bewertungen
- Alteração de Enzimas HepaticasDokument7 SeitenAlteração de Enzimas Hepaticasfernandojardim12100% (1)
- Proposta IDokument1 SeiteProposta Ifernandojardim12Noch keine Bewertungen
- Dislipidemias tratamentoDokument3 SeitenDislipidemias tratamentoAna Carolyna VargasNoch keine Bewertungen
- Bula PyloripacDokument59 SeitenBula Pyloripacfernandojardim12Noch keine Bewertungen
- Cap 9 - QUIMIOTERAPIA DO CANCER PDFDokument5 SeitenCap 9 - QUIMIOTERAPIA DO CANCER PDFfernandojardim12Noch keine Bewertungen
- Legislacao Brasileira Referente A RotulagemDokument11 SeitenLegislacao Brasileira Referente A RotulagemJuliana ZanetiNoch keine Bewertungen
- Comparação Entre Métodos Compostos No Cálculo de Afinidades Por Próton e ElétronDokument8 SeitenComparação Entre Métodos Compostos No Cálculo de Afinidades Por Próton e Elétronfernandojardim12Noch keine Bewertungen
- Aula - Descoberta e Desenvolvimento de FármacosDokument41 SeitenAula - Descoberta e Desenvolvimento de FármacosmarcosdiegoNoch keine Bewertungen
- Aula 03-QFIDokument3 SeitenAula 03-QFICecília CarvalhoNoch keine Bewertungen
- Guia de Iniciação e Configuração de App Inventor PB Revision 5Dokument59 SeitenGuia de Iniciação e Configuração de App Inventor PB Revision 5ElielNoch keine Bewertungen
- GROMACS Tutorial (Portuguese BR) v.1.6Dokument26 SeitenGROMACS Tutorial (Portuguese BR) v.1.6fernandojardim12Noch keine Bewertungen
- Descoberta de FarmacoDokument33 SeitenDescoberta de Farmacofernandojardim12Noch keine Bewertungen
- monoWMS MestradoDokument146 SeitenmonoWMS Mestradofernandojardim12Noch keine Bewertungen
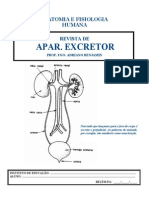
- REVISTA 8 Excretor 2010Dokument5 SeitenREVISTA 8 Excretor 2010fernandojardim12Noch keine Bewertungen
- Planejamento de FármacosDokument12 SeitenPlanejamento de FármacospollockpaguNoch keine Bewertungen
- TCC Opco 2Dokument10 SeitenTCC Opco 2fernandojardim12Noch keine Bewertungen
- Clister OpacoDokument1 SeiteClister Opacofernandojardim12Noch keine Bewertungen
- PE CB MM Fisiologia Geral Humana IDokument4 SeitenPE CB MM Fisiologia Geral Humana IWesley WinchesterNoch keine Bewertungen
- Avaliação Da Qualidade de Vida Dos Clientes Renais Crônicos em Tratamento HemodialiticoDokument98 SeitenAvaliação Da Qualidade de Vida Dos Clientes Renais Crônicos em Tratamento Hemodialiticofernandojardim12Noch keine Bewertungen
- PSORIASEDokument19 SeitenPSORIASEfernandojardim12Noch keine Bewertungen
- Cancer Colo RetalDokument37 SeitenCancer Colo Retalfernandojardim12Noch keine Bewertungen
- Apresentao Tcccleo01 12 2011 111201014331 Phpapp02Dokument22 SeitenApresentao Tcccleo01 12 2011 111201014331 Phpapp02fernandojardim12Noch keine Bewertungen
- Enema OpacoDokument25 SeitenEnema Opacofernandojardim12Noch keine Bewertungen
- Blefarite, Calázio e seu TratamentoDokument3 SeitenBlefarite, Calázio e seu Tratamentofernandojardim12Noch keine Bewertungen
- Instruções Sobre A Eloboração Do Artigo - Segunda ParteDokument3 SeitenInstruções Sobre A Eloboração Do Artigo - Segunda Partefernandojardim12Noch keine Bewertungen
- Livro Digital Conjuntivite ViralDokument19 SeitenLivro Digital Conjuntivite ViralSara KreboldNoch keine Bewertungen
- Poliomielite: a doença da paralisia infantilDokument19 SeitenPoliomielite: a doença da paralisia infantiljabire marijuvistaNoch keine Bewertungen
- 4ManualMensuraodeRiscoest avcolasdereproduoDSAFinalDokument16 Seiten4ManualMensuraodeRiscoest avcolasdereproduoDSAFinalMichelle GoulartNoch keine Bewertungen
- Instituto Industrial de MatundoDokument8 SeitenInstituto Industrial de MatundoMbolha Jose JequeNoch keine Bewertungen
- Lista Nacional de Notificação CompulsóriaDokument3 SeitenLista Nacional de Notificação CompulsóriaLucas SoaresNoch keine Bewertungen
- Universidade Federal de Santa Catarina Centro de Ciências Da Saúde Departamento de Análises Clínicas Curso de Graduação em FarmáciaDokument50 SeitenUniversidade Federal de Santa Catarina Centro de Ciências Da Saúde Departamento de Análises Clínicas Curso de Graduação em FarmáciaNanna Nail DesignerNoch keine Bewertungen
- Marcadores serológicos da hepatite BDokument1 SeiteMarcadores serológicos da hepatite BAna TeodoroNoch keine Bewertungen
- Fundamentos Da Microbiologia e ImunologiaDokument25 SeitenFundamentos Da Microbiologia e ImunologiaRaiane BritoNoch keine Bewertungen
- Resumo Do Filme ContágioDokument1 SeiteResumo Do Filme ContágioViviane InácioNoch keine Bewertungen
- Ensino Médio - Teste de Biologia com questões sobre recombinação genética, transferência de genes e agentes causadores de doençasDokument6 SeitenEnsino Médio - Teste de Biologia com questões sobre recombinação genética, transferência de genes e agentes causadores de doençasPaulo HenriqueNoch keine Bewertungen
- O que é a SIDA? Sintomas e como se previneDokument2 SeitenO que é a SIDA? Sintomas e como se previneStefanie Carey100% (1)
- Aula 2 - Aedes AegyptiDokument12 SeitenAula 2 - Aedes Aegyptiretrolipe08Noch keine Bewertungen
- Portugues Eleva Efai 4ano Cad2Dokument96 SeitenPortugues Eleva Efai 4ano Cad2amnykessiaNoch keine Bewertungen
- Recombinação genética e elementos móveis: exercícios de revisãoDokument3 SeitenRecombinação genética e elementos móveis: exercícios de revisãozycNoch keine Bewertungen
- Manual AEQ TRDokument18 SeitenManual AEQ TRChave pixNoch keine Bewertungen
- TópicosDokument41 SeitenTópicosngila88% (41)
- 1 Ano - BiologiaDokument3 Seiten1 Ano - BiologiaAnderson NovaisNoch keine Bewertungen
- Resolução de exercícios sobre habilidades de vidaDokument19 SeitenResolução de exercícios sobre habilidades de vidaVeronica Monteiro PedroNoch keine Bewertungen
- COVID-19 e coagulopatia: entenda os mecanismos da coagulopatia associadaDokument8 SeitenCOVID-19 e coagulopatia: entenda os mecanismos da coagulopatia associadaAndryellen Batista de CarvalhoNoch keine Bewertungen
- Infecção Pelo Vírus Da Imunodeficiência Humana (HIV) - Doenças Infecciosas - Manuais MSD Edição para ProfissionaisDokument30 SeitenInfecção Pelo Vírus Da Imunodeficiência Humana (HIV) - Doenças Infecciosas - Manuais MSD Edição para ProfissionaisIsaias MavuniceNoch keine Bewertungen
- Ppra Rocha e Dourado Aparecida de GoiâniaDokument62 SeitenPpra Rocha e Dourado Aparecida de GoiâniaWellington Barbosa BarrosNoch keine Bewertungen
- Trabalho Doenças Virais e Doenças BacterianasDokument9 SeitenTrabalho Doenças Virais e Doenças BacterianasMatheus Fai AlvesNoch keine Bewertungen
- Gabarito Prova PsicologiaDokument14 SeitenGabarito Prova PsicologiaGomes De LunaNoch keine Bewertungen
- 00 - Enem Por Assunto 2022 - Lásaro Henrique - Dom Barreto (Digital)Dokument165 Seiten00 - Enem Por Assunto 2022 - Lásaro Henrique - Dom Barreto (Digital)reboucasgiovana7Noch keine Bewertungen
- SES DF Enfermeiro - Revisão Doenças Transmissíveis, COVID e Epidemiologia Com Fernanda FeitosaDokument49 SeitenSES DF Enfermeiro - Revisão Doenças Transmissíveis, COVID e Epidemiologia Com Fernanda Feitosafabio sthuNoch keine Bewertungen
- Portefólio de Geografia C (Continuação)Dokument2 SeitenPortefólio de Geografia C (Continuação)Letícia SilvaNoch keine Bewertungen
- Doenças Exantemáticas - Frederico OásisDokument30 SeitenDoenças Exantemáticas - Frederico OásisRafael MedeirosNoch keine Bewertungen
- PBL 3º SemestreDokument15 SeitenPBL 3º SemestreLidia LizziNoch keine Bewertungen