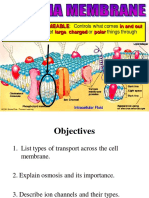
Das könnte Ihnen auch gefallen
- Biology: a QuickStudy Laminated Reference GuideVon EverandBiology: a QuickStudy Laminated Reference GuideBewertung: 3 von 5 Sternen3/5 (2)
- Assignment OneDokument6 SeitenAssignment OneEmmanuel OpokuNoch keine Bewertungen
- Active TransportDokument14 SeitenActive TransportRjay CruzNoch keine Bewertungen
- Membrane Transport Resource Notes - 2021Dokument36 SeitenMembrane Transport Resource Notes - 2021kxng crockedNoch keine Bewertungen
- Cell Membrane Function and StructureDokument6 SeitenCell Membrane Function and Structurecris ibarraNoch keine Bewertungen
- Cell Membrane TransportDokument5 SeitenCell Membrane Transportronsta86Noch keine Bewertungen
- Key Points RevisionDokument124 SeitenKey Points RevisionSumeyyah KemalNoch keine Bewertungen
- Aqa A Level Biology Importance of MembranesDokument3 SeitenAqa A Level Biology Importance of Membranesgayatribala306Noch keine Bewertungen
- Understanding Pathophysiology CH 1: Study Online atDokument6 SeitenUnderstanding Pathophysiology CH 1: Study Online atChristian del Rosario100% (2)
- The Role of Movement of Substances Across Membranes in The Function of Organs and Organ Systems.Dokument2 SeitenThe Role of Movement of Substances Across Membranes in The Function of Organs and Organ Systems.Bryan Earl Mwangi100% (6)
- 3 - kh201295 kh201295Dokument5 Seiten3 - kh201295 kh201295murtada gubaNoch keine Bewertungen
- Chapter 7 Membrane Structure and FunctionDokument4 SeitenChapter 7 Membrane Structure and FunctionMohammad Mahrus Bin AshfaqNoch keine Bewertungen
- Cellular ProcessesssssssssssssDokument10 SeitenCellular ProcessesssssssssssssJay Soledad ManganaNoch keine Bewertungen
- Test Essays January 4Dokument7 SeitenTest Essays January 4Stephanie JenkinsNoch keine Bewertungen
- 116 - Transport Mechanisms in CellsDokument4 Seiten116 - Transport Mechanisms in Cellslastjoe71100% (1)
- What Is Reverse OsmosisDokument7 SeitenWhat Is Reverse OsmosisAnggi AviandriNoch keine Bewertungen
- Active TransportDokument10 SeitenActive TransportSylvia DuzonNoch keine Bewertungen
- Chapter 5 Lesson 1-Cell Structures Involved in TransportDokument8 SeitenChapter 5 Lesson 1-Cell Structures Involved in TransportDaichi KatashiNoch keine Bewertungen
- Cell Membrane Grade 12Dokument8 SeitenCell Membrane Grade 12Olayinka SalmonNoch keine Bewertungen
- 2222 TransportDokument42 Seiten2222 TransportaddwrNoch keine Bewertungen
- Bio Notes FinalsDokument4 SeitenBio Notes FinalsJoaquine ArateaNoch keine Bewertungen
- Finals NotesDokument21 SeitenFinals NotesJoaquine ArateaNoch keine Bewertungen
- Biology (Free Response)Dokument6 SeitenBiology (Free Response)CynNoch keine Bewertungen
- Full Download Human Anatomy 4th Edition Saladin Solutions ManualDokument35 SeitenFull Download Human Anatomy 4th Edition Saladin Solutions Manualvocality.duettoqw603100% (36)
- Plasma Membrane: A Physical BarrierDokument3 SeitenPlasma Membrane: A Physical BarrierDiane OrsuaNoch keine Bewertungen
- Finals Bio 1 RevDokument6 SeitenFinals Bio 1 RevAnne Gelli ManaloNoch keine Bewertungen
- Prokaryotic and Eukaryotic CellsDokument6 SeitenProkaryotic and Eukaryotic CellsJam MoralejaNoch keine Bewertungen
- Human Anatomy 4th Edition Saladin Solutions ManualDokument25 SeitenHuman Anatomy 4th Edition Saladin Solutions ManualLeslieBuchanannjfy100% (52)
- CellDokument110 SeitenCellHeydrick GuyoNoch keine Bewertungen
- Notes For Summative Test 1Dokument67 SeitenNotes For Summative Test 1Phoemela Marie AlcaldeNoch keine Bewertungen
- 1 - Give An Illustrated Account of The Fine Staracture of A Plant Cell and Describe The Function of The Diffrent Cell OrgnagesDokument19 Seiten1 - Give An Illustrated Account of The Fine Staracture of A Plant Cell and Describe The Function of The Diffrent Cell OrgnagesMD shah KhanNoch keine Bewertungen
- Pys PDFDokument5 SeitenPys PDFMD shah KhanNoch keine Bewertungen
- Cells 3Dokument66 SeitenCells 3KEANNA RUBIANoch keine Bewertungen
- CellDokument112 SeitenCellHeydrick GuyoNoch keine Bewertungen
- Drug Absorption-WPS OfficeDokument6 SeitenDrug Absorption-WPS Officeloknath sur parthoNoch keine Bewertungen
- Physiology PPT Unit 1Dokument35 SeitenPhysiology PPT Unit 1vrindhasabu2004Noch keine Bewertungen
- Cytolog Membrtransport2 (Stud)Dokument4 SeitenCytolog Membrtransport2 (Stud)OnSolomonNoch keine Bewertungen
- General Biology 1 Module FINALSDokument12 SeitenGeneral Biology 1 Module FINALSKenneth Vince AlonzoNoch keine Bewertungen
- General Biology 1 Module FINALSDokument12 SeitenGeneral Biology 1 Module FINALSChristine Joyce Toledo VasquezNoch keine Bewertungen
- Note For Grade 10Dokument10 SeitenNote For Grade 10peter20000.npkNoch keine Bewertungen
- The Plasma Membrane: Membranes Cover The Surface of Every Cell, and Also Surround Most OrganellesDokument11 SeitenThe Plasma Membrane: Membranes Cover The Surface of Every Cell, and Also Surround Most OrganellesSyeda Kaneez Mohammadi MohammadiNoch keine Bewertungen
- Membrane Structure and FunctionDokument11 SeitenMembrane Structure and FunctionShiloh FrederickNoch keine Bewertungen
- Biology Study Guide (Wheaton) Complete)Dokument7 SeitenBiology Study Guide (Wheaton) Complete)Jireh HuangNoch keine Bewertungen
- Chapter 1 Cell Biology Lesson 2Dokument13 SeitenChapter 1 Cell Biology Lesson 2Juan GarciaNoch keine Bewertungen
- Cell MembraneDokument10 SeitenCell Membraneharshika tembhurneNoch keine Bewertungen
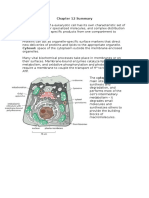
- Chapter 12 SummaryDokument7 SeitenChapter 12 SummaryCharlotteNoch keine Bewertungen
- Cell MembraneDokument15 SeitenCell MembraneAyesha SaleemNoch keine Bewertungen
- Electrical Gradient. An Equilibrium Is Reached at Which CL - Influx and CL - Efflux Are EqualDokument4 SeitenElectrical Gradient. An Equilibrium Is Reached at Which CL - Influx and CL - Efflux Are EqualMohib NirobNoch keine Bewertungen
- Cells: Transport of Materials Across The Cell MembraneDokument6 SeitenCells: Transport of Materials Across The Cell MembraneSara SalmanNoch keine Bewertungen
- Functions of The Cell MembraneDokument5 SeitenFunctions of The Cell Membranemekusa10Noch keine Bewertungen
- Mccance: Pathophysiology, 6Th Edition: Chapter 01: Cellular Biology Key Points - Print Summary ReviewDokument5 SeitenMccance: Pathophysiology, 6Th Edition: Chapter 01: Cellular Biology Key Points - Print Summary ReviewAbiah IsraelNoch keine Bewertungen
- Cell Le1Dokument14 SeitenCell Le1eng.raghdah98Noch keine Bewertungen
- ASSIGMENT of Environmental Toxicology 12Dokument8 SeitenASSIGMENT of Environmental Toxicology 12naeem attaNoch keine Bewertungen
- N-Linked Protein Glycosylation Occurs in All Three Life-Form Domains: Eukaryotes, Bacteria, andDokument9 SeitenN-Linked Protein Glycosylation Occurs in All Three Life-Form Domains: Eukaryotes, Bacteria, andHeisenbergNoch keine Bewertungen
- Chapter 2 Student Copy ZoologyDokument5 SeitenChapter 2 Student Copy Zoologyapi-523871804Noch keine Bewertungen
- Cell Structure and Function: Tenets of Cell TheoryDokument9 SeitenCell Structure and Function: Tenets of Cell Theoryjey011Noch keine Bewertungen
- General Physiology of Excitable Tissues: Lecture in Normal PhysiologyDokument54 SeitenGeneral Physiology of Excitable Tissues: Lecture in Normal Physiologymohammad farooque razaaNoch keine Bewertungen
- Cell ConstituentsDokument8 SeitenCell ConstituentsSanaa SamkoNoch keine Bewertungen
- Achilike Sbi Assignment 1Dokument6 SeitenAchilike Sbi Assignment 1skillsj123Noch keine Bewertungen
- Antivirals For HerpesDokument4 SeitenAntivirals For HerpesBrandon BarndtNoch keine Bewertungen
- Enzymes ChartDokument8 SeitenEnzymes ChartBrandon BarndtNoch keine Bewertungen
- Glycolysis and The TCA Cycle: Glycogen PathwaysDokument8 SeitenGlycolysis and The TCA Cycle: Glycogen PathwaysBrandon BarndtNoch keine Bewertungen
- 01 Introduction HandoutDokument14 Seiten01 Introduction HandoutBrandon BarndtNoch keine Bewertungen
- What Becomes What DevelopmentDokument10 SeitenWhat Becomes What DevelopmentBrandon BarndtNoch keine Bewertungen
- Bio I Master GuideDokument44 SeitenBio I Master GuideBrandon BarndtNoch keine Bewertungen
- Resistive Circuit PracticeDokument11 SeitenResistive Circuit PracticeBrandon BarndtNoch keine Bewertungen
- EssayDokument10 SeitenEssayBrandon BarndtNoch keine Bewertungen
- Price ListDokument28 SeitenPrice ListBrandon BarndtNoch keine Bewertungen
- STR Mpa-MpmDokument8 SeitenSTR Mpa-MpmBANGGANoch keine Bewertungen
- Endothermic Gas Production Overview: Tmosphere Ngineering OmpanyDokument6 SeitenEndothermic Gas Production Overview: Tmosphere Ngineering OmpanyJhon ChitNoch keine Bewertungen
- Onuaguluchi1996 1Dokument10 SeitenOnuaguluchi1996 1IkaSugihartatikNoch keine Bewertungen
- BS 65-1981Dokument27 SeitenBS 65-1981jasonNoch keine Bewertungen
- New Book "101 Costly HR Mistakes... and How To Fix Them" by Vanessa Nelson Released To Help Employers Avoid Costly HR Mistakes and Save MillionsDokument2 SeitenNew Book "101 Costly HR Mistakes... and How To Fix Them" by Vanessa Nelson Released To Help Employers Avoid Costly HR Mistakes and Save MillionsPR.comNoch keine Bewertungen
- My Public Self My Hidden Self My Blind Spots My Unknown SelfDokument2 SeitenMy Public Self My Hidden Self My Blind Spots My Unknown SelfMaria Hosanna PalorNoch keine Bewertungen
- Bitumen BasicsDokument25 SeitenBitumen BasicsMILON KUMAR HORENoch keine Bewertungen
- Boeco BM-800 - User ManualDokument21 SeitenBoeco BM-800 - User ManualJuan Carlos CrespoNoch keine Bewertungen
- Unknown Facts About Physicians Email List - AverickMediaDokument13 SeitenUnknown Facts About Physicians Email List - AverickMediaJames AndersonNoch keine Bewertungen
- Radproduction Chapter 2-9Dokument276 SeitenRadproduction Chapter 2-9Christian DioNoch keine Bewertungen
- Gay Costa Del Sol - 2010Dokument2 SeitenGay Costa Del Sol - 2010gayinfospainNoch keine Bewertungen
- Maya Mendez ResumeDokument2 SeitenMaya Mendez Resumeapi-520985654Noch keine Bewertungen
- Cemco T80Dokument140 SeitenCemco T80Eduardo Ariel Bernal100% (3)
- Challenger 350 Recommended Operating Procedures and TechniquesDokument104 SeitenChallenger 350 Recommended Operating Procedures and Techniquessebatsea100% (1)
- WeaknessesDokument4 SeitenWeaknessesshyamiliNoch keine Bewertungen
- Chapter 4 CrystallizationDokument13 SeitenChapter 4 Crystallizationprosedur0% (1)
- Derivative Investment!Dokument24 SeitenDerivative Investment!Asif Riaz100% (2)
- Business PlanDokument17 SeitenBusiness PlanChester Cortez50% (2)
- ClistDokument14 SeitenClistGuerraNoch keine Bewertungen
- Tetra Pak Training CatalogueDokument342 SeitenTetra Pak Training CatalogueElif UsluNoch keine Bewertungen
- Chapter 7 Unemployment, Inflation, and Long-Run GrowthDokument21 SeitenChapter 7 Unemployment, Inflation, and Long-Run GrowthNataly FarahNoch keine Bewertungen
- Datasheet MEC MPS200 v1 2018Dokument4 SeitenDatasheet MEC MPS200 v1 2018Cepi Sindang KamulanNoch keine Bewertungen
- Material Safety Data Sheet: Wonder Gel™ Stainless Steel Pickling GelDokument2 SeitenMaterial Safety Data Sheet: Wonder Gel™ Stainless Steel Pickling GelTrần Thùy LinhNoch keine Bewertungen
- Total Elbow Arthroplasty and RehabilitationDokument5 SeitenTotal Elbow Arthroplasty and RehabilitationMarina ENoch keine Bewertungen
- Scoop of Practice aOTADokument9 SeitenScoop of Practice aOTAfercespedNoch keine Bewertungen
- Geography - Development (Rural - Urban Settlement)Dokument32 SeitenGeography - Development (Rural - Urban Settlement)jasmine le rouxNoch keine Bewertungen
- Constantino V MendezDokument3 SeitenConstantino V MendezNīc CādīgālNoch keine Bewertungen
- Aromatic Electrophilic SubstitutionDokument71 SeitenAromatic Electrophilic SubstitutionsridharancNoch keine Bewertungen
- Case Report 3 MukokelDokument3 SeitenCase Report 3 MukokelWidychii GadiestchhetyaNoch keine Bewertungen
- Universal ING - LA.Boschi Plants Private LimitedDokument23 SeitenUniversal ING - LA.Boschi Plants Private LimitedAlvaro Mendoza MaytaNoch keine Bewertungen