Das könnte Ihnen auch gefallen
- 2021 DesignDokument20 Seiten2021 Designbdvd1007092Noch keine Bewertungen
- Genetic Design 2021Dokument33 SeitenGenetic Design 2021bdvd1007092Noch keine Bewertungen
- Steel Joint Consortium Iran Afghanistan Russia ChiDokument20 SeitenSteel Joint Consortium Iran Afghanistan Russia Chibdvd1007092Noch keine Bewertungen
- Childhood and Adolescent Cancer Statistics, 2014 PDFDokument21 SeitenChildhood and Adolescent Cancer Statistics, 2014 PDFErlitaa LarasatiNoch keine Bewertungen
- Self-Assembly of DNA Molecules Towards DNA NanorobDokument3 SeitenSelf-Assembly of DNA Molecules Towards DNA Nanorobbdvd1007092Noch keine Bewertungen
- Critical Review of The TransCelerate Template ForDokument10 SeitenCritical Review of The TransCelerate Template Forbdvd1007092Noch keine Bewertungen
- Feasibility and Reliability of Sequential Logic With Gene HALDokument20 SeitenFeasibility and Reliability of Sequential Logic With Gene HALbdvd1007092Noch keine Bewertungen
- Synthetic Biology Christopher A VoigtDokument2 SeitenSynthetic Biology Christopher A Voigtbdvd1007092Noch keine Bewertungen
- The EMBO Journal - 2020 - Lukassen - SARS CoV 2 Receptor ACE2 and TMPRSS2 Are Primarily Expressed in Bronchial TransientDokument15 SeitenThe EMBO Journal - 2020 - Lukassen - SARS CoV 2 Receptor ACE2 and TMPRSS2 Are Primarily Expressed in Bronchial Transientbdvd1007092Noch keine Bewertungen
- Valuation of RD Compound Option Using Markov ChainDokument27 SeitenValuation of RD Compound Option Using Markov Chainbdvd1007092Noch keine Bewertungen
- 2 - 1 Koriyama - MultivariableDokument36 Seiten2 - 1 Koriyama - Multivariablebdvd1007092Noch keine Bewertungen
- Ama-Style - FinalDokument35 SeitenAma-Style - Finalbdvd1007092Noch keine Bewertungen
- Multivariate Regression ModellingDokument10 SeitenMultivariate Regression Modellingbdvd1007092Noch keine Bewertungen
- Multivariate Regression ModellingDokument10 SeitenMultivariate Regression Modellingbdvd1007092Noch keine Bewertungen
- Fgene 05 00132Dokument6 SeitenFgene 05 00132bdvd1007092Noch keine Bewertungen
- Ni Hms 583013Dokument18 SeitenNi Hms 583013bdvd1007092Noch keine Bewertungen
- Bmri2014 512831Dokument14 SeitenBmri2014 512831bdvd1007092Noch keine Bewertungen
- 1475 925X 13 64Dokument17 Seiten1475 925X 13 64bdvd1007092Noch keine Bewertungen
- Indoleamine 2,3-Dioxygenase (IDO) Is Frequently Expressed in Stromal Cells of Hodgkin Lymphoma and Is Associated With Adverse Clinical Features: A Retrospective Cohort StudyDokument9 SeitenIndoleamine 2,3-Dioxygenase (IDO) Is Frequently Expressed in Stromal Cells of Hodgkin Lymphoma and Is Associated With Adverse Clinical Features: A Retrospective Cohort Studybdvd1007092Noch keine Bewertungen
- NIH Public Access: Adaptive TherapyDokument22 SeitenNIH Public Access: Adaptive Therapybdvd1007092Noch keine Bewertungen
- Blood Pressure and Flow Ch10Dokument20 SeitenBlood Pressure and Flow Ch10bdvd1007092Noch keine Bewertungen
- NIH Public Access: Author ManuscriptDokument13 SeitenNIH Public Access: Author Manuscriptbdvd1007092Noch keine Bewertungen
- nm0514 454Dokument2 Seitennm0514 454bdvd1007092Noch keine Bewertungen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (119)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Venipuncture Techniques and ProceduresDokument66 SeitenVenipuncture Techniques and ProceduresAngelica Camille B. AbaoNoch keine Bewertungen
- AntihypertensivesDokument35 SeitenAntihypertensivesjanuajipatriaNoch keine Bewertungen
- Auxiliary VerbsDokument12 SeitenAuxiliary VerbsNoe Lia CastroNoch keine Bewertungen
- Pediatrics ImpDokument233 SeitenPediatrics Impملك عيسى100% (1)
- Blood Supply of The Head, Neck and BrainDokument34 SeitenBlood Supply of The Head, Neck and BrainChong Xue FengNoch keine Bewertungen
- Science Magazine, Issue 6657 (August 4, 2023)Dokument175 SeitenScience Magazine, Issue 6657 (August 4, 2023)Kim LevrelNoch keine Bewertungen
- Introduction of PHCDokument39 SeitenIntroduction of PHCIdiris Mohamed100% (1)
- Cement CSRDokument35 SeitenCement CSRKasak Gupta100% (1)
- Elective Caesarean Section on a GoatDokument17 SeitenElective Caesarean Section on a GoatAbdirazak AlkhaalidNoch keine Bewertungen
- Male infertility evaluation: what you need to knowDokument1 SeiteMale infertility evaluation: what you need to knowa jNoch keine Bewertungen
- SEMINAR-Clinical Microbiology: Topic-Human Immuno-Deficiency Virus (HIV)Dokument54 SeitenSEMINAR-Clinical Microbiology: Topic-Human Immuno-Deficiency Virus (HIV)sushant_dharNoch keine Bewertungen
- Essential Health Services and PlansDokument4 SeitenEssential Health Services and Planszahara mahalNoch keine Bewertungen
- Renal Vegetarian NutritionDokument2 SeitenRenal Vegetarian NutritionSg Balaji100% (1)
- Trilobata Exhibits Molecular Impediment On TheDokument10 SeitenTrilobata Exhibits Molecular Impediment On TheInternational Journal of Innovative Science and Research TechnologyNoch keine Bewertungen
- Nursing Diagnosis For AsthmaDokument6 SeitenNursing Diagnosis For AsthmaTINAIDA33% (3)
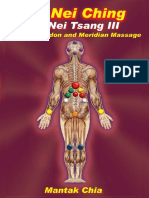
- BL42-Chi Nei Tsang III (ID-PDF) 12-17-15 PDFDokument90 SeitenBL42-Chi Nei Tsang III (ID-PDF) 12-17-15 PDFJosé Tao75% (8)
- Arthur Adolphus Lindsay - Mind The Transformer The New Psychology Complete (1911)Dokument172 SeitenArthur Adolphus Lindsay - Mind The Transformer The New Psychology Complete (1911)truepotentialzNoch keine Bewertungen
- NASAL SEPTUM DEVIATION: CAUSES, SYMPTOMS AND SURGICAL CORRECTIONDokument105 SeitenNASAL SEPTUM DEVIATION: CAUSES, SYMPTOMS AND SURGICAL CORRECTIONNguyễn ThànhNoch keine Bewertungen
- Imaging of Nasopharyngeal CarcinomaDokument9 SeitenImaging of Nasopharyngeal CarcinomayohanasmjtNoch keine Bewertungen
- Research Proposal First Draft (Adamu Moti)Dokument25 SeitenResearch Proposal First Draft (Adamu Moti)ODAA TUBENoch keine Bewertungen
- BSDokument6 SeitenBSBeda MalecdanNoch keine Bewertungen
- Fall 2010 Eco Newsletter, EcoSuperiorDokument12 SeitenFall 2010 Eco Newsletter, EcoSuperiorEco SuperiorNoch keine Bewertungen
- Diabetic Ketoacidosis in PregnancyDokument12 SeitenDiabetic Ketoacidosis in PregnancyMuhammad BilalNoch keine Bewertungen
- Iet Me Introduce My Name Najib MahfuzhDokument2 SeitenIet Me Introduce My Name Najib MahfuzhnajibNoch keine Bewertungen
- Biological Science Reviewer QuestionsDokument14 SeitenBiological Science Reviewer QuestionsRavian Mhe BitonNoch keine Bewertungen
- Health Promotion and Disease Prevention in the ElderlyDokument1 SeiteHealth Promotion and Disease Prevention in the ElderlyBeverly PagcaliwaganNoch keine Bewertungen
- Congenital SyphilisDokument28 SeitenCongenital SyphilisMeena Koushal100% (4)
- GEHC Brochure Senographe CareDokument7 SeitenGEHC Brochure Senographe CareVremedSoluCionesNoch keine Bewertungen
- ZZZZZZZZZZZZZZDokument8 SeitenZZZZZZZZZZZZZZchloramphenicolNoch keine Bewertungen