Das könnte Ihnen auch gefallen
- Histeria Por Vacunación Mass Psychogenic Illness After Vaccination 2003Dokument6 SeitenHisteria Por Vacunación Mass Psychogenic Illness After Vaccination 2003Oscar Alejandro Cardenas QuinteroNoch keine Bewertungen
- Depresion y Ejercicio NejmDokument2 SeitenDepresion y Ejercicio NejmOscar Alejandro Cardenas QuinteroNoch keine Bewertungen
- Depresion PostpartoDokument6 SeitenDepresion PostpartoOscar Alejandro Cardenas QuinteroNoch keine Bewertungen
- Depresion en El Anciano en APS NejmDokument3 SeitenDepresion en El Anciano en APS NejmOscar Alejandro Cardenas QuinteroNoch keine Bewertungen
- Depression As An Evolutionary Strategy For Defense Against InfectionDokument14 SeitenDepression As An Evolutionary Strategy For Defense Against InfectionOscar Alejandro Cardenas Quintero100% (2)
- Differential Typology of Delusions in Major Depression and Schizophrenia. A Critique To The Unitary Concept of PsychosisDokument8 SeitenDifferential Typology of Delusions in Major Depression and Schizophrenia. A Critique To The Unitary Concept of PsychosisOscar Alejandro Cardenas QuinteroNoch keine Bewertungen
- Cjp-June-05 - GEDokument2 SeitenCjp-June-05 - GEOscar Alejandro Cardenas QuinteroNoch keine Bewertungen
- V71I12 - A Breakthrough in Schizophrenia GeneticsDokument2 SeitenV71I12 - A Breakthrough in Schizophrenia GeneticsOscar Alejandro Cardenas QuinteroNoch keine Bewertungen
- Diagnosis and Clinical Features of Major Neuropsychiatric Disorders in HIV InfectionDokument9 SeitenDiagnosis and Clinical Features of Major Neuropsychiatric Disorders in HIV InfectionOscar Alejandro Cardenas QuinteroNoch keine Bewertungen
- A Case of Nonconvulsive Status Epilepticus Presenting As Dissociative FugueDokument2 SeitenA Case of Nonconvulsive Status Epilepticus Presenting As Dissociative FugueOscar Alejandro Cardenas QuinteroNoch keine Bewertungen
- Kapur (2003) Psychosis As A State of Aberrant SalienceDokument11 SeitenKapur (2003) Psychosis As A State of Aberrant SalienceOscar Alejandro Cardenas QuinteroNoch keine Bewertungen
- Kapur (2003) Psychosis As A State of Aberrant SalienceDokument11 SeitenKapur (2003) Psychosis As A State of Aberrant SalienceOscar Alejandro Cardenas QuinteroNoch keine Bewertungen
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5795)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Erin Lommen - Adrenal Fatigue or Sagging SeratoninDokument18 SeitenErin Lommen - Adrenal Fatigue or Sagging Seratoningreencrystal81Noch keine Bewertungen
- Questions & Answers: For For For For For NEET (UG) - 2020Dokument21 SeitenQuestions & Answers: For For For For For NEET (UG) - 2020Ritu JoharNoch keine Bewertungen
- EMG Methods For Evaluating Muscle and Nerve FunctionDokument546 SeitenEMG Methods For Evaluating Muscle and Nerve Function4569077100% (1)
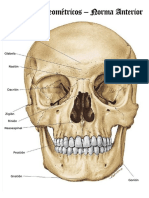
- Puntos Craneometricos CompressDokument13 SeitenPuntos Craneometricos CompressLaura PNoch keine Bewertungen
- Canine and Feline Endocrinology PDFDokument685 SeitenCanine and Feline Endocrinology PDFSandra Solano100% (8)
- Lecture Notes On Biophysics and Bioimaging 2009Dokument128 SeitenLecture Notes On Biophysics and Bioimaging 2009Anonymous 9rJe2lOskx100% (2)
- Bowel Nosodes Power PointDokument15 SeitenBowel Nosodes Power PointTheresa Dale83% (6)
- Human Body EssayDokument1 SeiteHuman Body EssayAtlas CruxNoch keine Bewertungen
- CamScanner 10-29-2020 13.11.47Dokument69 SeitenCamScanner 10-29-2020 13.11.47Yuki ChanNoch keine Bewertungen
- Shoku AnafilaktikDokument16 SeitenShoku AnafilaktikindeenikeNoch keine Bewertungen
- SLPDB PCN 1999Dokument3 SeitenSLPDB PCN 1999KSI ShawonNoch keine Bewertungen
- Vital SignsDokument3 SeitenVital SignsNicole NipasNoch keine Bewertungen
- Newport, D.J. and Nemeroff, C.B. Neurobiology of Posttraumatic Stress Disorder. Cognitive Neuroscience JAARTALDokument8 SeitenNewport, D.J. and Nemeroff, C.B. Neurobiology of Posttraumatic Stress Disorder. Cognitive Neuroscience JAARTALg10564433Noch keine Bewertungen
- Best HST WorkoutDokument29 SeitenBest HST WorkoutVova ShushNoch keine Bewertungen
- Status EpilepticusDokument70 SeitenStatus Epilepticusfloppyfishh100% (1)
- Canine and Feline Endocrinology and ReproductionDokument1.095 SeitenCanine and Feline Endocrinology and ReproductionmarparolaNoch keine Bewertungen
- Cerebrovascular Disease (Emphasis On CVA)Dokument29 SeitenCerebrovascular Disease (Emphasis On CVA)OlynsieMorrisNoch keine Bewertungen
- Intro To Ana Physio RBLDokument69 SeitenIntro To Ana Physio RBLHades HadesNoch keine Bewertungen
- II. Blood Vessels Physiology By: Dr. Abdulrahman Aqra MD MSCDokument12 SeitenII. Blood Vessels Physiology By: Dr. Abdulrahman Aqra MD MSCapi-290338101Noch keine Bewertungen
- Airway Management - IntubationDokument31 SeitenAirway Management - IntubationRajaNoch keine Bewertungen
- Summer Training For Speed: Alwyn Cosgrove's Training Design ProgramDokument6 SeitenSummer Training For Speed: Alwyn Cosgrove's Training Design ProgramSteve HeywoodNoch keine Bewertungen
- ADR Common Drug - Induced Organ DisordersDokument45 SeitenADR Common Drug - Induced Organ Disorders0009439Noch keine Bewertungen
- Effect of Plant Growth Enhancers On Growth and Flowering of Tuberose Cv. Prajwal PDFDokument4 SeitenEffect of Plant Growth Enhancers On Growth and Flowering of Tuberose Cv. Prajwal PDFSupal DesaiNoch keine Bewertungen
- Effective Suction-Imporession For Retentive Mand Dentture PDFDokument24 SeitenEffective Suction-Imporession For Retentive Mand Dentture PDFFaheemuddin MuhammadNoch keine Bewertungen
- The Powerful Impact of StressDokument19 SeitenThe Powerful Impact of StressMichael Angelo SeñaNoch keine Bewertungen
- 11 Ch1208 5127 PDFDokument25 Seiten11 Ch1208 5127 PDFThamarai ElanthirayanNoch keine Bewertungen
- Kami Export - Patricia Isabel Sandoval Martínez - Circ. Syst. ST - ANSWER SHEET 12-26Dokument4 SeitenKami Export - Patricia Isabel Sandoval Martínez - Circ. Syst. ST - ANSWER SHEET 12-26p.sandoval.2027Noch keine Bewertungen
- Emma Jordan - Research Final DraftDokument7 SeitenEmma Jordan - Research Final Draftapi-436829682Noch keine Bewertungen
- U.S. Army Survival Manual FM 21-76 (June 1992)Dokument572 SeitenU.S. Army Survival Manual FM 21-76 (June 1992)explorer_ebook100% (1)
- Antioxidants !! The Great DiscoveryDokument3 SeitenAntioxidants !! The Great DiscoverypatNoch keine Bewertungen