Das könnte Ihnen auch gefallen
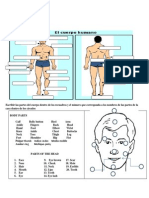
- Tamil Names For Water BodiesDokument3 SeitenTamil Names For Water BodiesINDIRAKALYANI100% (2)
- Are Human Beings Impossibe To ApeDokument4 SeitenAre Human Beings Impossibe To ApeINDIRAKALYANINoch keine Bewertungen
- Coronal Hole in SunDokument2 SeitenCoronal Hole in SunINDIRAKALYANINoch keine Bewertungen
- Nutritional Value Per 100 G (3.5 Oz) : EnergyDokument3 SeitenNutritional Value Per 100 G (3.5 Oz) : EnergyINDIRAKALYANINoch keine Bewertungen
- Features Extracted From The Energy Conservation Act, 2001Dokument1 SeiteFeatures Extracted From The Energy Conservation Act, 2001INDIRAKALYANINoch keine Bewertungen
- Top 5 As On 16.10.2012Dokument7 SeitenTop 5 As On 16.10.2012INDIRAKALYANINoch keine Bewertungen
- Monitoring Patients With M ProteinDokument1 SeiteMonitoring Patients With M ProteinINDIRAKALYANINoch keine Bewertungen
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Exam TrematodeDokument16 SeitenExam Trematodelamiazak52100% (2)
- DNA TribesDokument20 SeitenDNA TribeskomooryNoch keine Bewertungen
- Chimie Spécialisée - Hormone Profil Diabetique: Analyse Resultats Unite Valeur NormaleDokument6 SeitenChimie Spécialisée - Hormone Profil Diabetique: Analyse Resultats Unite Valeur NormaleRajeev PareekNoch keine Bewertungen
- Lynn Saxon - The Naked Bonobo (2016)Dokument142 SeitenLynn Saxon - The Naked Bonobo (2016)jooo93100% (1)
- MCQ Question BankDokument102 SeitenMCQ Question BankdrmadaanpiyushNoch keine Bewertungen
- Science 6 Quiz 1 & 2, QTR 2Dokument2 SeitenScience 6 Quiz 1 & 2, QTR 2Reymart B Tiosan100% (1)
- RHS Plants RHS Tall Bearded Iris TrialDokument2 SeitenRHS Plants RHS Tall Bearded Iris TrialCebotari TatianaNoch keine Bewertungen
- Anatomical TerminologyDokument12 SeitenAnatomical Terminologysarped10Noch keine Bewertungen
- InglesDokument46 SeitenInglesAzulit KryNoch keine Bewertungen
- Artistic Anatomy of The Human F - Henry WarrenDokument51 SeitenArtistic Anatomy of The Human F - Henry WarrenGerman Enrique MartinezNoch keine Bewertungen
- THE SHIFTER'S KISS Erotic Paranormal Romance NovellaDokument8 SeitenTHE SHIFTER'S KISS Erotic Paranormal Romance NovellaCaridad Pineiro100% (1)
- Matary Surgical Anatomy 2013Dokument320 SeitenMatary Surgical Anatomy 2013Raouf Ra'fat Soliman96% (23)
- Triptico BocaDokument2 SeitenTriptico BocaRosa Muñoz100% (1)
- PercyDokument17 SeitenPercyRohit David Joseph0% (1)
- What Is Nervous Tissue?: Nerve Tissue Are All Made Up of SpecialisedDokument17 SeitenWhat Is Nervous Tissue?: Nerve Tissue Are All Made Up of SpecialisedAtyyahEsirNoch keine Bewertungen
- 10 Simple Home Remedies For Flawless Smooth Glowing SkinDokument2 Seiten10 Simple Home Remedies For Flawless Smooth Glowing SkinFirdous BhatNoch keine Bewertungen
- A Plea For AnimalsDokument3 SeitenA Plea For AnimalsdsolisNoch keine Bewertungen
- 23 Anatomy Physiology TermsDokument30 Seiten23 Anatomy Physiology TermsAlyssa Gwyneth GalubaNoch keine Bewertungen
- Bertalanffy Ludwig Von 1957 Quantitative Laws in Metabolism and GrowthDokument16 SeitenBertalanffy Ludwig Von 1957 Quantitative Laws in Metabolism and GrowthCarlos AndradeNoch keine Bewertungen
- Alice Wade - Flame ThrowerDokument191 SeitenAlice Wade - Flame ThrowerAnca BuzeaNoch keine Bewertungen
- Module (Notes) Science Form 2Dokument48 SeitenModule (Notes) Science Form 2Nur Hafizah Alwi40% (5)
- 4-Active and Retentive Components ofDokument16 Seiten4-Active and Retentive Components ofKhalid MortajaNoch keine Bewertungen
- Reproduction in PlantsDokument14 SeitenReproduction in PlantsAsif AyazNoch keine Bewertungen
- Junyoung ParkDokument2 SeitenJunyoung Parkapi-295865543Noch keine Bewertungen
- Herodians Family TreeDokument1 SeiteHerodians Family TreeKelan JenkinsNoch keine Bewertungen
- Mirror Neurons and The Evolution of LanguageDokument11 SeitenMirror Neurons and The Evolution of LanguagejkmilNoch keine Bewertungen
- Histology of Naval CavityDokument66 SeitenHistology of Naval CavityRomaine-Ricardo FrancisNoch keine Bewertungen
- Bent Over RowDokument3 SeitenBent Over Rowserveh80Noch keine Bewertungen
- CHPT 42 Circulation Gas Exchange 2014Dokument32 SeitenCHPT 42 Circulation Gas Exchange 2014azn2017Noch keine Bewertungen
- Structure of The Teeth and Supporting Tissues: Reading Assignment: Afp 162-6, Vol. 1, Pages 84 - 85Dokument22 SeitenStructure of The Teeth and Supporting Tissues: Reading Assignment: Afp 162-6, Vol. 1, Pages 84 - 85harnageaNoch keine Bewertungen