Das könnte Ihnen auch gefallen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Emergency Care Algorithms 2019 PDFDokument75 SeitenEmergency Care Algorithms 2019 PDFsiraj ammed75% (4)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- RCDSO Guidelines Implant Dentistry PDFDokument16 SeitenRCDSO Guidelines Implant Dentistry PDFl4j0b9Noch keine Bewertungen
- OsteoarthritisDokument4 SeitenOsteoarthritisDharti PatelNoch keine Bewertungen
- Design Calc - Cooling Tower Sizing - R1 - 15.10.2011Dokument1 SeiteDesign Calc - Cooling Tower Sizing - R1 - 15.10.2011AliAliNoch keine Bewertungen
- Laboratory Test InterpretationDokument73 SeitenLaboratory Test Interpretationjosieangel11Noch keine Bewertungen
- Interpretation MRCPCH 2009 Site: For Part 2 Exam For Part 2 ExamDokument35 SeitenInterpretation MRCPCH 2009 Site: For Part 2 Exam For Part 2 Examyassine100% (1)
- Water, Sanitation and Hygiene Manual: WASH Training For Hygiene Promotion StaffDokument20 SeitenWater, Sanitation and Hygiene Manual: WASH Training For Hygiene Promotion StaffSadashiv RahaneNoch keine Bewertungen
- Fpsne 100Dokument1 SeiteFpsne 100AliAliNoch keine Bewertungen
- Water Treatment - Overview Ion ExchangeDokument9 SeitenWater Treatment - Overview Ion ExchangeAliAliNoch keine Bewertungen
- Tuberculosis (TB) Prevention and Control Workplace Policy and ProgramDokument3 SeitenTuberculosis (TB) Prevention and Control Workplace Policy and ProgramluckyNoch keine Bewertungen
- Immunoassay For Estrogens in The Environment: Multi-Analyte - Detektion Based On FluorescenceDokument1 SeiteImmunoassay For Estrogens in The Environment: Multi-Analyte - Detektion Based On FluorescenceAliAliNoch keine Bewertungen
- Anakon 2001Dokument1 SeiteAnakon 2001AliAliNoch keine Bewertungen
- Exercice 1 Peptide Mass Fingerprinting Exercise (Mass List)Dokument3 SeitenExercice 1 Peptide Mass Fingerprinting Exercise (Mass List)AliAliNoch keine Bewertungen
- Preface About The Author: Wastes BODDokument6 SeitenPreface About The Author: Wastes BODAliAliNoch keine Bewertungen
- Front Matter1Dokument1 SeiteFront Matter1AliAliNoch keine Bewertungen
- In Our 2009-2010 Project, We Have Successfully Developed A Novel Method Which Combines ImmunomagneticDokument2 SeitenIn Our 2009-2010 Project, We Have Successfully Developed A Novel Method Which Combines ImmunomagneticAliAliNoch keine Bewertungen
- Index: Index Terms Links ADokument11 SeitenIndex: Index Terms Links AAliAliNoch keine Bewertungen
- FPSBa 8cDokument1 SeiteFPSBa 8cAliAliNoch keine Bewertungen
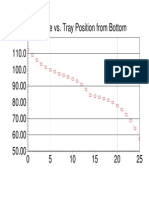
- 120.0 Temperature vs. Tray Position From BottomDokument1 Seite120.0 Temperature vs. Tray Position From BottomAliAliNoch keine Bewertungen
- Material Stream: F-G: ConditionsDokument2 SeitenMaterial Stream: F-G: ConditionsAliAliNoch keine Bewertungen
- Important Compatability Superlasticizes Portland CementDokument23 SeitenImportant Compatability Superlasticizes Portland CementAliAliNoch keine Bewertungen
- FPSXL 2LDokument1 SeiteFPSXL 2LAliAliNoch keine Bewertungen
- Fps Magna Cide 575Dokument1 SeiteFps Magna Cide 575AliAliNoch keine Bewertungen
- FPSMS 90Dokument1 SeiteFPSMS 90AliAliNoch keine Bewertungen
- Drink Water On Empty StomachDokument4 SeitenDrink Water On Empty StomachbabuNoch keine Bewertungen
- Test Bank For College Algebra 12Th Edition by Lial Hornsby Schneider and Daniels Isbn 0134217454 978013421745 Full Chapter PDFDokument36 SeitenTest Bank For College Algebra 12Th Edition by Lial Hornsby Schneider and Daniels Isbn 0134217454 978013421745 Full Chapter PDFdavid.weldon592100% (12)
- Hemophilia and Its Treatment: Brief ReviewDokument7 SeitenHemophilia and Its Treatment: Brief ReviewSalsa BillaNoch keine Bewertungen
- BodyDokument124 SeitenBodygion.nandNoch keine Bewertungen
- 3aph Plant InfoDokument27 Seiten3aph Plant InfoPatricia De AsisNoch keine Bewertungen
- 3 - Physical Examination of UrineDokument6 Seiten3 - Physical Examination of UrineKunware TropaNoch keine Bewertungen
- Annex A - Self-Assessment Tool For Proposed EPCB Health Facilities PDFDokument5 SeitenAnnex A - Self-Assessment Tool For Proposed EPCB Health Facilities PDFCora Mendoza100% (1)
- Aedes Egypti ADokument10 SeitenAedes Egypti AOswin YohsaNoch keine Bewertungen
- Distinct Spectrum of CFTR Gene MutationsDokument13 SeitenDistinct Spectrum of CFTR Gene Mutationsapi-3742014Noch keine Bewertungen
- SAMPLE Outline PanicDokument3 SeitenSAMPLE Outline PanicdexNoch keine Bewertungen
- Intracardiac Epinephrine Injection During Open Thoracotomy and Circulatory Arrest 2155 6148.1000341Dokument2 SeitenIntracardiac Epinephrine Injection During Open Thoracotomy and Circulatory Arrest 2155 6148.1000341samuelNoch keine Bewertungen
- Report 1 - History of Medical TechnologyDokument1 SeiteReport 1 - History of Medical TechnologyMaxine TaeyeonNoch keine Bewertungen
- Chapter 20 Extrahepatic Biliary Tract and Gall BladderDokument63 SeitenChapter 20 Extrahepatic Biliary Tract and Gall BladderMACON824Noch keine Bewertungen
- The Science of Sleep: A Brief Guide On How To Sleep Better Every NightDokument2 SeitenThe Science of Sleep: A Brief Guide On How To Sleep Better Every NightMark Anthony RaymundoNoch keine Bewertungen
- Rural Awareness Work Experience Programme Manual: B.Sc. (Hons.) HorticultureDokument88 SeitenRural Awareness Work Experience Programme Manual: B.Sc. (Hons.) HorticulturePotsangbam Dhanabir SinghNoch keine Bewertungen
- Abreu, CarlosDokument1 SeiteAbreu, CarlosCarlos AbreuNoch keine Bewertungen
- Project ENGLISH 2019 BacteriaDokument4 SeitenProject ENGLISH 2019 BacteriaAna Maria MacoveiNoch keine Bewertungen
- Clinical Manifestations and Assessment of Respiratory Disease 5th Edition Jardins Test BankDokument9 SeitenClinical Manifestations and Assessment of Respiratory Disease 5th Edition Jardins Test Banksarahpalmerotpdkjcwfq100% (26)
- Manual Physical Therapy Certificate Program: CurriculumDokument8 SeitenManual Physical Therapy Certificate Program: CurriculumAsim SayyadNoch keine Bewertungen
- Sensititre Plate Guide Booklet EN PDFDokument62 SeitenSensititre Plate Guide Booklet EN PDFXuân Tài NguyễnNoch keine Bewertungen
- EpidemiologiDokument7 SeitenEpidemiologiazzaNoch keine Bewertungen
- FORMULASI ANTISEPTIK TANGAN EKSTRAK DAUN SIRIH (Piper Betle L.) DENGAN BAHAN PENSTABIL TEA (Trietanolamin)Dokument9 SeitenFORMULASI ANTISEPTIK TANGAN EKSTRAK DAUN SIRIH (Piper Betle L.) DENGAN BAHAN PENSTABIL TEA (Trietanolamin)nirmala evelynNoch keine Bewertungen
- PCORI Methodology Standards Curriculum Research Questions 3Dokument13 SeitenPCORI Methodology Standards Curriculum Research Questions 3Krizele Acu PagalananNoch keine Bewertungen