Das könnte Ihnen auch gefallen
- Effect of Spatial Orientation of The Horizontal Semicircular Canal On The Vestibulo-Ocular ReflexDokument8 SeitenEffect of Spatial Orientation of The Horizontal Semicircular Canal On The Vestibulo-Ocular ReflexRudolfGerNoch keine Bewertungen
- Analysis of Persistent Geotropic and Apogeotropic Positional Nystagmus of The Lateral Canal Benign Paroxysmal Positional VertigoDokument6 SeitenAnalysis of Persistent Geotropic and Apogeotropic Positional Nystagmus of The Lateral Canal Benign Paroxysmal Positional VertigoRudolfGerNoch keine Bewertungen
- VNG CB en Rev02 - HDDokument5 SeitenVNG CB en Rev02 - HDRudolfGerNoch keine Bewertungen
- Abnormal Cancellation of The Vestibulo-Ocular ReflexDokument4 SeitenAbnormal Cancellation of The Vestibulo-Ocular ReflexRudolfGerNoch keine Bewertungen
- A Simple Gain-Based Evaluation of The Video Head Impulse Test Reliably Detects Normal Vestibulo-Ocular Reflex Indicative of Stroke in Patients With Acute Vestibular SyndromeDokument8 SeitenA Simple Gain-Based Evaluation of The Video Head Impulse Test Reliably Detects Normal Vestibulo-Ocular Reflex Indicative of Stroke in Patients With Acute Vestibular SyndromeRudolfGerNoch keine Bewertungen
- COVID-19 and Compulsory Vaccination - AnDokument24 SeitenCOVID-19 and Compulsory Vaccination - AnRudolfGerNoch keine Bewertungen
- Latest Statistics On England Mortality Data Suggest Systematic Mis-Categorisation of Vaccine Status and Uncertain Effectiveness of Covid-19Dokument24 SeitenLatest Statistics On England Mortality Data Suggest Systematic Mis-Categorisation of Vaccine Status and Uncertain Effectiveness of Covid-19RudolfGerNoch keine Bewertungen
- Concussion Guidelines Step 2 - Evidence For Subtype ClassificationDokument12 SeitenConcussion Guidelines Step 2 - Evidence For Subtype ClassificationRudolfGerNoch keine Bewertungen
- Head-Jolting Nystagmus Occlusion of The Horizontal Semicircular Canal Induced by Vigorous Head ShakingDokument4 SeitenHead-Jolting Nystagmus Occlusion of The Horizontal Semicircular Canal Induced by Vigorous Head ShakingRudolfGerNoch keine Bewertungen
- Causes and Characteristics of Horizontal Positional NystagmusDokument9 SeitenCauses and Characteristics of Horizontal Positional NystagmusRudolfGerNoch keine Bewertungen
- Pelvic Pain With Sitting: Diagnostic Algorithm: A. Lee Dellon, MD, PHD, Johns Hopkins UniversityDokument1 SeitePelvic Pain With Sitting: Diagnostic Algorithm: A. Lee Dellon, MD, PHD, Johns Hopkins UniversityRudolfGerNoch keine Bewertungen
- Fall Prediction in Neurological Gait Disorders DifDokument15 SeitenFall Prediction in Neurological Gait Disorders DifRudolfGerNoch keine Bewertungen
- The Spectrum of Vestibular MigraineDokument14 SeitenThe Spectrum of Vestibular MigraineRudolfGerNoch keine Bewertungen
- A Pilot Study To Evaluate Multi-Dimensional Effects of Dance For People With Parkinson's DiseaseDokument6 SeitenA Pilot Study To Evaluate Multi-Dimensional Effects of Dance For People With Parkinson's DiseaseRudolfGerNoch keine Bewertungen
- Cervical Spine Pre-Screening For Arterial DysfunctionDokument11 SeitenCervical Spine Pre-Screening For Arterial DysfunctionRudolfGerNoch keine Bewertungen
- The Treatment of Chronic Coccydynia With PDFDokument7 SeitenThe Treatment of Chronic Coccydynia With PDFRudolfGerNoch keine Bewertungen
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- 40 Sa Hindi - LBDokument6 Seiten40 Sa Hindi - LBIrshad mohammedNoch keine Bewertungen
- Whitepaper Psychographic SegmentationDokument32 SeitenWhitepaper Psychographic SegmentationAubreyNoch keine Bewertungen
- Karvonen Formula Worksheet PDFDokument2 SeitenKarvonen Formula Worksheet PDFpipninjaNoch keine Bewertungen
- Peer ReviewDokument24 SeitenPeer ReviewSimon Dzokoto100% (1)
- V Jttu/: Prices ChangeDokument179 SeitenV Jttu/: Prices ChangeKarpincho3Noch keine Bewertungen
- Icare Pivc GuidelineDokument26 SeitenIcare Pivc GuidelineCarissa CkNoch keine Bewertungen
- Ebook Healing PrayersDokument12 SeitenEbook Healing PrayersAllen Bethea100% (1)
- 2010 Endocrine System ChartDokument3 Seiten2010 Endocrine System ChartAlexander QianNoch keine Bewertungen
- Guiding Principles For Best Practices in Geriatric PTDokument16 SeitenGuiding Principles For Best Practices in Geriatric PTPritesh KujurNoch keine Bewertungen
- 95 Formulation and Evaluation of Diclofenac Sodium Gel by Using Natural PolymerDokument3 Seiten95 Formulation and Evaluation of Diclofenac Sodium Gel by Using Natural PolymerJulian Kayne100% (1)
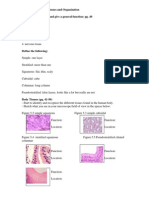
- Human Biology Laboratory 4: Body Tissues and Organization List The 4 Tissue Types and Give A General Function: Pg. 40Dokument4 SeitenHuman Biology Laboratory 4: Body Tissues and Organization List The 4 Tissue Types and Give A General Function: Pg. 40bugyourselfNoch keine Bewertungen
- Makalah Bahasa InggrisDokument15 SeitenMakalah Bahasa Inggrisfera kartikaNoch keine Bewertungen
- What Is Personal HygieneDokument4 SeitenWhat Is Personal HygieneAnonymous 0FWhoTu100% (1)
- A Review On Aleurites MoluccanaDokument18 SeitenA Review On Aleurites Moluccanabharatkhurana85Noch keine Bewertungen
- Heart Attack & Drinking Warm WaterDokument1 SeiteHeart Attack & Drinking Warm Waterjoshua_sx180% (5)
- Farm ChemicalsDokument16 SeitenFarm ChemicalsHasrul RosliNoch keine Bewertungen
- Order Denying NHL Motion To DismissDokument33 SeitenOrder Denying NHL Motion To DismissNHL Concussion LawsuitNoch keine Bewertungen
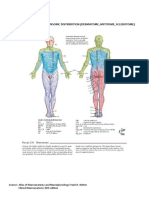
- Dermatome Myotome SclerotomeDokument4 SeitenDermatome Myotome SclerotomeAdhya TiaraNoch keine Bewertungen
- Transcript of Sun Pharma Conference Call On Halol FacilityDokument20 SeitenTranscript of Sun Pharma Conference Call On Halol FacilityNed PagliaruloNoch keine Bewertungen
- Drug Education NotesDokument75 SeitenDrug Education NotesJoshua D None-NoneNoch keine Bewertungen
- Overview He Most As IsDokument20 SeitenOverview He Most As IsManushi ChauhanNoch keine Bewertungen
- Fitzgerald AnnInternMed 1999 130 70 Curiosity PDFDokument3 SeitenFitzgerald AnnInternMed 1999 130 70 Curiosity PDFPaulinaNoch keine Bewertungen
- IonnyDokument4 SeitenIonnyLu KaNoch keine Bewertungen
- Chinese Medicine PsychologyDokument315 SeitenChinese Medicine PsychologyPaco Ladera100% (5)
- Big RPLDokument1 SeiteBig RPLRahayu LestariNoch keine Bewertungen
- Fluids and Electrolytes Study GuideDokument13 SeitenFluids and Electrolytes Study GuideElizabeth McKeeNoch keine Bewertungen
- Chapter - 014 Student HandoutsDokument16 SeitenChapter - 014 Student Handoutsebiniyam2021Noch keine Bewertungen
- Raw Materials & Purification - NitrogenDokument9 SeitenRaw Materials & Purification - Nitrogensexyrusty0% (1)
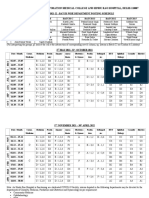
- Interns 2016 DistributionDokument13 SeitenInterns 2016 DistributionKashish ChawlaNoch keine Bewertungen
- Psychiatric HistoryDokument63 SeitenPsychiatric HistoryStarlet Rhonadez Bito-onon Oriel0% (1)