Das könnte Ihnen auch gefallen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Lexus GS300 Wiring DiagramDokument4 SeitenLexus GS300 Wiring Diagramrenz100% (1)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- EncoperesisDokument6 SeitenEncoperesisRishi SelvamNoch keine Bewertungen
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Basic Statistics in Bio MedicalDokument271 SeitenBasic Statistics in Bio Medicalifai.gs7487100% (2)
- Guidelines TAVI TAVRDokument15 SeitenGuidelines TAVI TAVRRossellaDisalvo100% (1)
- Theory of HumoursDokument19 SeitenTheory of HumoursSyed Ahad0% (1)
- Ayurveda FitoterapiaDokument112 SeitenAyurveda FitoterapiaKerol Bomfim100% (2)
- The Impact of Tumor Biology On Cancer Treatment and Multidisciplinary Strategies - M. Molls, Et Al., (Springer, 2009) WWDokument363 SeitenThe Impact of Tumor Biology On Cancer Treatment and Multidisciplinary Strategies - M. Molls, Et Al., (Springer, 2009) WWiuliNoch keine Bewertungen
- Case Study - Seizuring DogDokument8 SeitenCase Study - Seizuring Dogapi-301746262Noch keine Bewertungen
- AONE Nursing CompetenciesDokument11 SeitenAONE Nursing CompetenciesAmi NoviantiNoch keine Bewertungen
- Immunolocalization of Aquaporin-5 Expression in Sweat Gland Cells From Normal and Anhidrotic Horses (Pages 17-23)Dokument7 SeitenImmunolocalization of Aquaporin-5 Expression in Sweat Gland Cells From Normal and Anhidrotic Horses (Pages 17-23)jenNoch keine Bewertungen
- Art Theraphy AutismDokument2 SeitenArt Theraphy AutismMatheaFiliNoch keine Bewertungen
- Spondyloarthritis ResidentDokument70 SeitenSpondyloarthritis Residentviraaj pawar100% (1)
- Feline Plasma Cell Pododermatitis - A Study of 8 Cases (Pages 333-337) PDFDokument5 SeitenFeline Plasma Cell Pododermatitis - A Study of 8 Cases (Pages 333-337) PDFjenNoch keine Bewertungen
- Effects of Dietary Flax Seed and Sunflower Seed Supplementation On Normal Canine Serum Polyunsaturated Fatty Acids and Skin and Hair PDFDokument7 SeitenEffects of Dietary Flax Seed and Sunflower Seed Supplementation On Normal Canine Serum Polyunsaturated Fatty Acids and Skin and Hair PDFjenNoch keine Bewertungen
- Isolation of Microsporum Canis From The Hair Coat of Pet Dogs and Cats Belonging To Owners Diagnosed With M. Canis Tinea Corporis (Pages 327-331)Dokument5 SeitenIsolation of Microsporum Canis From The Hair Coat of Pet Dogs and Cats Belonging To Owners Diagnosed With M. Canis Tinea Corporis (Pages 327-331)jenNoch keine Bewertungen
- Oestrogen Receptor Evaluation in Pomeranian Dogs With Hair Cycle Arrest (Alopecia X) On Melatonin Supplementation (Pages 252-258)Dokument7 SeitenOestrogen Receptor Evaluation in Pomeranian Dogs With Hair Cycle Arrest (Alopecia X) On Melatonin Supplementation (Pages 252-258)jenNoch keine Bewertungen
- Patch Testing of Experimentally Sensitized Beagle Dogs - Development of A Model For Skin Lesions of Atopic Dermatitis (Pages 95-102)Dokument8 SeitenPatch Testing of Experimentally Sensitized Beagle Dogs - Development of A Model For Skin Lesions of Atopic Dermatitis (Pages 95-102)jenNoch keine Bewertungen
- Mouse Epidermal Development - Effects of Retinoic Acid Exposure in Utero (Pages 36-44)Dokument9 SeitenMouse Epidermal Development - Effects of Retinoic Acid Exposure in Utero (Pages 36-44)jenNoch keine Bewertungen
- The Impact of Body Site, Topical Melatonin and Brushing On Hair Regrowth After Clipping Normal Siberian Husky Dogs (Pages 45-50)Dokument6 SeitenThe Impact of Body Site, Topical Melatonin and Brushing On Hair Regrowth After Clipping Normal Siberian Husky Dogs (Pages 45-50)jenNoch keine Bewertungen
- A Retrospective Study of Cutaneous Equine Sarcoidosis and Its Potential Infectious Aetiological Agents (Pages 51-62)Dokument12 SeitenA Retrospective Study of Cutaneous Equine Sarcoidosis and Its Potential Infectious Aetiological Agents (Pages 51-62)jenNoch keine Bewertungen
- Generalized Calcinosis Cutis Associated With Probable Leptospirosis in A Dog (Pages 401-406)Dokument6 SeitenGeneralized Calcinosis Cutis Associated With Probable Leptospirosis in A Dog (Pages 401-406)jenNoch keine Bewertungen
- The Pathophysiology of Otosclerosis: Review of Current ResearchDokument6 SeitenThe Pathophysiology of Otosclerosis: Review of Current ResearchFelipeAsenjoÁlvarezNoch keine Bewertungen
- 4 - Chronic InfectionsDokument55 Seiten4 - Chronic InfectionsAlexa GabrielaNoch keine Bewertungen
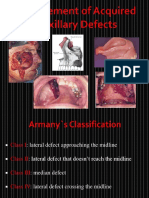
- Management of Acquired Maxillary Defects Partially Edentlous PDFDokument129 SeitenManagement of Acquired Maxillary Defects Partially Edentlous PDFRamy Khalifa AshmawyNoch keine Bewertungen
- Suvarnprashan The Concept of Ayurveda Immunisation: Pharma Science Monitor 7 (4), Oct-Dec 2016Dokument5 SeitenSuvarnprashan The Concept of Ayurveda Immunisation: Pharma Science Monitor 7 (4), Oct-Dec 2016AtulSanapNoch keine Bewertungen
- Comparing Different Approaches To CounsellingDokument7 SeitenComparing Different Approaches To CounsellingTim RichardsonNoch keine Bewertungen
- Arixtra: (Fondaparinux Sodium) InjectionDokument26 SeitenArixtra: (Fondaparinux Sodium) InjectionTri Purma SariNoch keine Bewertungen
- Simultaneous Determination of Methylparaben + Propylparaben + Hidrocortisone Topical Cream PDFDokument7 SeitenSimultaneous Determination of Methylparaben + Propylparaben + Hidrocortisone Topical Cream PDFNájla KassabNoch keine Bewertungen
- Rational Use of The Drug (Rud) : Rovina Ruslami, DR., SPPD, PHDDokument31 SeitenRational Use of The Drug (Rud) : Rovina Ruslami, DR., SPPD, PHDamaliaramadhaniNoch keine Bewertungen
- Seizure Updated ILAE ClassificationDokument12 SeitenSeizure Updated ILAE ClassificationNasheei RadjaNoch keine Bewertungen
- G.R. No. 132319. May 12, 2000Dokument2 SeitenG.R. No. 132319. May 12, 2000Chua RoxyNoch keine Bewertungen
- This Study Resource WasDokument3 SeitenThis Study Resource WasCarlito AglipayNoch keine Bewertungen
- Assessment of The Use of A Dental Mouth MirrorDokument7 SeitenAssessment of The Use of A Dental Mouth MirrorIOSRjournalNoch keine Bewertungen
- Pharamcology Unit 10 AnswersDokument13 SeitenPharamcology Unit 10 AnswersLovren YoungNoch keine Bewertungen
- Doctor's OrderDokument2 SeitenDoctor's OrderFranzky HernandezNoch keine Bewertungen
- Hospital and Clinical Pharmacist PDFDokument23 SeitenHospital and Clinical Pharmacist PDFmajd67% (6)
- False Advertisement Gluten Free FoodDokument9 SeitenFalse Advertisement Gluten Free FoodElenaNoch keine Bewertungen
- PREP & COGENT - 13th and 14th December 2014Dokument6 SeitenPREP & COGENT - 13th and 14th December 2014FdghNoch keine Bewertungen
- 05 2019 Pediatric Infectious Diseases LAYOUT R3Dokument58 Seiten05 2019 Pediatric Infectious Diseases LAYOUT R3khalid alharbiNoch keine Bewertungen