Das könnte Ihnen auch gefallen
- Development of A Standardized Set of Patient-Centered Outcomes For Advanced Prostate CancerDokument8 SeitenDevelopment of A Standardized Set of Patient-Centered Outcomes For Advanced Prostate CanceritontNoch keine Bewertungen
- United Theory of Planet Formation (I) - Tandem RegimeDokument17 SeitenUnited Theory of Planet Formation (I) - Tandem RegimeitontNoch keine Bewertungen
- Oclc Worldshare Collection Manager Quick Start Guide: Initial Sign-In / Change PasswordDokument14 SeitenOclc Worldshare Collection Manager Quick Start Guide: Initial Sign-In / Change PassworditontNoch keine Bewertungen
- Self-Organizing Systems in Planetary Physics: Harmonic Resonances of Planet and Moon OrbitsDokument17 SeitenSelf-Organizing Systems in Planetary Physics: Harmonic Resonances of Planet and Moon OrbitsitontNoch keine Bewertungen
- Probabilistic Modelling of Security of Supply in Gas Networks and Evaluation of New InfrastructureDokument11 SeitenProbabilistic Modelling of Security of Supply in Gas Networks and Evaluation of New InfrastructureitontNoch keine Bewertungen
- Mining Software Usage With The Automatic Library TrackingDokument10 SeitenMining Software Usage With The Automatic Library TrackingitontNoch keine Bewertungen
- The Ethics of Marketing CancerDokument2 SeitenThe Ethics of Marketing CanceritontNoch keine Bewertungen
- Mining Software Usage With The Automatic Library TrackingDokument10 SeitenMining Software Usage With The Automatic Library TrackingitontNoch keine Bewertungen
- The Multi-Objective Generalized Consistent Vehicle Routing ProblemDokument18 SeitenThe Multi-Objective Generalized Consistent Vehicle Routing ProblemitontNoch keine Bewertungen
- Place-Keeping in Action: Evaluating The Capacity of Green Space Partnerships in EnglandDokument11 SeitenPlace-Keeping in Action: Evaluating The Capacity of Green Space Partnerships in EnglanditontNoch keine Bewertungen
- Metabolic Approach To Heart Failure - The Role Og Metabolic ModulatorsDokument5 SeitenMetabolic Approach To Heart Failure - The Role Og Metabolic ModulatorsitontNoch keine Bewertungen
- Estimation of The Critical Velocity in Pipeline Flow of SlurriesDokument13 SeitenEstimation of The Critical Velocity in Pipeline Flow of Slurriesitont100% (1)
- Prostate CancerDokument8 SeitenProstate CanceritontNoch keine Bewertungen
- Validation Theory and The Myth of The TherapeuticDokument2 SeitenValidation Theory and The Myth of The TherapeuticitontNoch keine Bewertungen
- Seminars in Cancer BiologyDokument9 SeitenSeminars in Cancer BiologyitontNoch keine Bewertungen
- 1 s2.0 S1877782114002239 MainDokument7 Seiten1 s2.0 S1877782114002239 MainitontNoch keine Bewertungen
- Seminars in Cancer BiologyDokument9 SeitenSeminars in Cancer BiologyitontNoch keine Bewertungen
- Effects of Second and Subsequent LinesDokument8 SeitenEffects of Second and Subsequent LinesitontNoch keine Bewertungen
- Combining Immunotherapy and Radiation For Prostate Cancer: ReviewDokument9 SeitenCombining Immunotherapy and Radiation For Prostate Cancer: ReviewitontNoch keine Bewertungen
- 1 s2.0 S2213596014001111 MainDokument4 Seiten1 s2.0 S2213596014001111 MainitontNoch keine Bewertungen
- Theoretical Considerations of RES-Avoiding LiposomesDokument20 SeitenTheoretical Considerations of RES-Avoiding LiposomesitontNoch keine Bewertungen
- 1 s2.0 S0959804914010582 MainDokument11 Seiten1 s2.0 S0959804914010582 MainitontNoch keine Bewertungen
- Journal of Atmospheric Solar-Terrestrial PhysicsDokument8 SeitenJournal of Atmospheric Solar-Terrestrial PhysicsitontNoch keine Bewertungen
- Seminars in Cancer BiologyDokument9 SeitenSeminars in Cancer BiologyitontNoch keine Bewertungen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (120)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Epilepsi 1Dokument12 SeitenEpilepsi 1Ima SoniaNoch keine Bewertungen
- Coااببmplete denture handoutsDokument18 SeitenCoااببmplete denture handoutsطارق حمادةNoch keine Bewertungen
- Obat Induksi HIPERURISEMIA A PDFDokument10 SeitenObat Induksi HIPERURISEMIA A PDFshelfinararaNoch keine Bewertungen
- Considerations For The Design of Antibody-Based TherapeuticsDokument30 SeitenConsiderations For The Design of Antibody-Based TherapeuticsDaniel CastanNoch keine Bewertungen
- Charles DissertationDokument81 SeitenCharles DissertationMhlanga Bridget TanyaradzwaNoch keine Bewertungen
- The Effect of Omega-3 Fatty Acids On Active Periodontal Therapy: A Systematic Review and Meta-AnalysisDokument14 SeitenThe Effect of Omega-3 Fatty Acids On Active Periodontal Therapy: A Systematic Review and Meta-AnalysisDadi SindhuNoch keine Bewertungen
- Task 2 Engineering English (Redho Triwinoko, METO 5B)Dokument3 SeitenTask 2 Engineering English (Redho Triwinoko, METO 5B)Redho TriwinokoNoch keine Bewertungen
- Laboratory Exercise No. 10 Endocrine SystemDokument3 SeitenLaboratory Exercise No. 10 Endocrine SystemJamesanne DemetriaNoch keine Bewertungen
- Dr. Nugroho Sigit SpRad PDFDokument20 SeitenDr. Nugroho Sigit SpRad PDFMira HandayaniNoch keine Bewertungen
- Journal .Of Orthodontics and Oral Surgery: AmericanDokument11 SeitenJournal .Of Orthodontics and Oral Surgery: AmericanShirley AliagaNoch keine Bewertungen
- Tiered Response AgreementDokument3 SeitenTiered Response AgreementThe Hamilton SpectatorNoch keine Bewertungen
- Hypertension LecturesDokument65 SeitenHypertension LecturesAdebisiNoch keine Bewertungen
- Maternal Health Research Priorities PDFDokument9 SeitenMaternal Health Research Priorities PDFIshrat PatelNoch keine Bewertungen
- Rupture UterusDokument20 SeitenRupture UterusGOODE ALBERTNoch keine Bewertungen
- Cebu City Medical Center-College of NursingDokument5 SeitenCebu City Medical Center-College of NursingJimnah Rhodrick BontilaoNoch keine Bewertungen
- Ardc 2Dokument1 SeiteArdc 2Camille GuintoNoch keine Bewertungen
- Team Work EssayDokument7 SeitenTeam Work Essayezknbk5h100% (2)
- 2015 Nature Glypican 1 Identifies Cancer ExosomesDokument24 Seiten2015 Nature Glypican 1 Identifies Cancer ExosomesAgata HugglerNoch keine Bewertungen
- PSI INDIA - Will Balbir Pasha Help Fight AIDS: Target AudienceDokument2 SeitenPSI INDIA - Will Balbir Pasha Help Fight AIDS: Target AudienceShambhawi SinhaNoch keine Bewertungen
- Lovejoy 1985Dokument10 SeitenLovejoy 1985mikulcicaNoch keine Bewertungen
- Dermawound Original Venous Stasis Wound CareDokument8 SeitenDermawound Original Venous Stasis Wound CareNew Medical Solutions - Your Source for Dermawound, BurnBGone and more...Noch keine Bewertungen
- Resmed Astral 150Dokument93 SeitenResmed Astral 150Lê Văn KhánhNoch keine Bewertungen
- Preprint Not Peer Reviewed: COVID-19 and Lockdown: A Study On The Impact On Mental HealthDokument13 SeitenPreprint Not Peer Reviewed: COVID-19 and Lockdown: A Study On The Impact On Mental HealthloloasbNoch keine Bewertungen
- 0529 Protocol Update 6.2.09 PDFDokument57 Seiten0529 Protocol Update 6.2.09 PDFTowhidulIslamNoch keine Bewertungen
- Patient Education Power Point DVTDokument14 SeitenPatient Education Power Point DVTMihaela PopescuNoch keine Bewertungen
- 2014 Group Case StudyDokument42 Seiten2014 Group Case StudyBrian100% (1)
- The Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition (DSM-5)Dokument42 SeitenThe Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition (DSM-5)Mihaela Onisia UngureanuNoch keine Bewertungen
- SirkulasiDokument17 SeitenSirkulasiJessica VanyaNoch keine Bewertungen
- Adnexal Torsion: Clinical, Radiological and Pathological Characteristics in A Tertiary Care Centre in Southern IndiaDokument6 SeitenAdnexal Torsion: Clinical, Radiological and Pathological Characteristics in A Tertiary Care Centre in Southern IndiaKriti KumariNoch keine Bewertungen
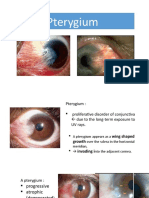
- 2020 - PterygiumDokument11 Seiten2020 - PterygiumDimas Agung Oko PutraNoch keine Bewertungen