Das könnte Ihnen auch gefallen
- Sex HormonesDokument58 SeitenSex Hormonesbug4444not100% (1)
- How To Read A CTGDokument12 SeitenHow To Read A CTGyehezkieldward100% (1)
- Ovulation Induction Evidence Based GuidelinesDokument207 SeitenOvulation Induction Evidence Based Guidelineswaleedali100% (3)
- Reproductive Endocrinology FinalDokument98 SeitenReproductive Endocrinology FinalChino Paolo SamsonNoch keine Bewertungen
- Nutr Influences EstrogenDokument8 SeitenNutr Influences EstrogenPortiaI100% (1)
- 2008 AJHG ZhuDokument8 Seiten2008 AJHG ZhuHaojie ZhuNoch keine Bewertungen
- Assignment On Obstetrics and Gynaecology DepartmentDokument14 SeitenAssignment On Obstetrics and Gynaecology Departmentbhavnil_1796Noch keine Bewertungen
- OBS & GYN MCQsDokument10 SeitenOBS & GYN MCQsmaximNoch keine Bewertungen
- ObGyn Outline BeckmannDokument85 SeitenObGyn Outline Beckmannp4sierra50% (2)
- Melatonine 3Dokument16 SeitenMelatonine 3unrealragdollsNoch keine Bewertungen
- Case Summary Ovarian CystDokument4 SeitenCase Summary Ovarian CystFrancesca AkanjiNoch keine Bewertungen
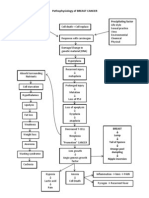
- Pathophysiology of BREAST CANCERDokument1 SeitePathophysiology of BREAST CANCERAlinor Abubacar100% (6)
- Lp1 ReviewerDokument17 SeitenLp1 ReviewerNiña Ricci MtflcoNoch keine Bewertungen
- Lepidium Meyenii (Maca), A Root With Aphrodisiac andDokument6 SeitenLepidium Meyenii (Maca), A Root With Aphrodisiac anda303111Noch keine Bewertungen
- Lepidium Meyenii (Maca), A Root With Aphrodisiac andDokument6 SeitenLepidium Meyenii (Maca), A Root With Aphrodisiac andDavid DizonNoch keine Bewertungen
- Physiological Effects of Melatonin On Leptin, Testosterone and Biochemical Parameters in Albino RatsDokument6 SeitenPhysiological Effects of Melatonin On Leptin, Testosterone and Biochemical Parameters in Albino RatsIOSR Journal of PharmacyNoch keine Bewertungen
- Male Axillary ExtractsDokument7 SeitenMale Axillary ExtractsbeemitsuNoch keine Bewertungen
- Impact of Alcohol On Male Reproductive Hormones Oxidative Stress and Semen Parameters in Sprague Dawley RatsDokument5 SeitenImpact of Alcohol On Male Reproductive Hormones Oxidative Stress and Semen Parameters in Sprague Dawley RatsSophia SantosNoch keine Bewertungen
- Che Cura 2010Dokument8 SeitenChe Cura 2010camila andrea rivera diazNoch keine Bewertungen
- Karatsoreos Et Al 2010Dokument11 SeitenKaratsoreos Et Al 2010zacharymweil100% (1)
- Studying The Role of Leptinand Adiponectin Hormones and Their Relation With Some Reproductive Hormones in Breastfeeding WomenDokument9 SeitenStudying The Role of Leptinand Adiponectin Hormones and Their Relation With Some Reproductive Hormones in Breastfeeding WomenadraaNoch keine Bewertungen
- Purwoceng Dan AfrodisiakDokument10 SeitenPurwoceng Dan AfrodisiakAnonymous HPmfOqdwNoch keine Bewertungen
- Original Article: Key Words: Bisphenol A, Luteinizing HormoneDokument6 SeitenOriginal Article: Key Words: Bisphenol A, Luteinizing HormoneAhmedkhaedNoch keine Bewertungen
- The Effect of Celery (Apium Graveolens) Extract On The Reproductive Hormones in Male MiceDokument6 SeitenThe Effect of Celery (Apium Graveolens) Extract On The Reproductive Hormones in Male MiceMihaela CiochinăNoch keine Bewertungen
- Orexin A and B Levels in The Hypothalamus of Female Rats: The Effects of The Estrous Cycle and AgeDokument6 SeitenOrexin A and B Levels in The Hypothalamus of Female Rats: The Effects of The Estrous Cycle and AgeNeha WanareNoch keine Bewertungen
- NIH Public Access: Author ManuscriptDokument24 SeitenNIH Public Access: Author ManuscriptnikuNoch keine Bewertungen
- 1 s2.0 S0196978108001320 MainDokument7 Seiten1 s2.0 S0196978108001320 MainTeodora OnofreiNoch keine Bewertungen
- RodrguezFuentes G - Effect of Hyperosmotic 2009Dokument4 SeitenRodrguezFuentes G - Effect of Hyperosmotic 2009Gabriela RodriguezNoch keine Bewertungen
- ECAM2015 324369 PDFDokument6 SeitenECAM2015 324369 PDFLuiz Otavio CostaNoch keine Bewertungen
- 4 Effect of Protein and Calorie Malnutrition On Drug MetabolismDokument11 Seiten4 Effect of Protein and Calorie Malnutrition On Drug MetabolismAdip100% (1)
- 17-Estradiol Attenuates Hippocampal Neuronal LossDokument9 Seiten17-Estradiol Attenuates Hippocampal Neuronal LossrodrigounitedNoch keine Bewertungen
- The Effect of Opiates On The Activity of Human Placental Aromatase CYP19 (2007)Dokument8 SeitenThe Effect of Opiates On The Activity of Human Placental Aromatase CYP19 (2007)Nimra Naveed ShaikhNoch keine Bewertungen
- The Hypothalamus-Pituitary-Ovary Axis and Type 1 Diabetes Mellitus: A Mini ReviewDokument11 SeitenThe Hypothalamus-Pituitary-Ovary Axis and Type 1 Diabetes Mellitus: A Mini ReviewdrmaruliNoch keine Bewertungen
- The Inhibitory Effect of Anandamide On Luteinizing Hormone-Releasing Hormone Secretion Is Reversed by EstrogenDokument6 SeitenThe Inhibitory Effect of Anandamide On Luteinizing Hormone-Releasing Hormone Secretion Is Reversed by EstrogenVeronica TestaNoch keine Bewertungen
- Effects of Acute Prolactin Manipulation On Sexual Drive and Function in MalesDokument9 SeitenEffects of Acute Prolactin Manipulation On Sexual Drive and Function in MalesAudi GohNoch keine Bewertungen
- Femael Infertility PDFDokument9 SeitenFemael Infertility PDFAyadPalaniNoch keine Bewertungen
- Michela Marinelli Et Al - Glucocorticoids and Behavioral Effects of Psychostimulants. II: Cocaine Intravenous Self-Administration and Reinstatement Depend On Glucocorticoid LevelsDokument7 SeitenMichela Marinelli Et Al - Glucocorticoids and Behavioral Effects of Psychostimulants. II: Cocaine Intravenous Self-Administration and Reinstatement Depend On Glucocorticoid LevelsSour60Noch keine Bewertungen
- Hormone of Reproductive System (Synthesis and Mechanism of Action)Dokument38 SeitenHormone of Reproductive System (Synthesis and Mechanism of Action)pwfNoch keine Bewertungen
- BIOLOGY OF REPRODUCTION 50, 765-773 (1994) : 'SupportedDokument9 SeitenBIOLOGY OF REPRODUCTION 50, 765-773 (1994) : 'SupportedLateecka R KulkarniNoch keine Bewertungen
- Animal Models For Human Polycystic Ovary Syndrome (PCOS) Focused On The Use of Indirect Hormonal Perturbations: A Review of The LiteratureDokument27 SeitenAnimal Models For Human Polycystic Ovary Syndrome (PCOS) Focused On The Use of Indirect Hormonal Perturbations: A Review of The LiteratureSamantha Orozco PinedaNoch keine Bewertungen
- Black Cohosh: Insights Into Its Mechanism(s) of ActionDokument12 SeitenBlack Cohosh: Insights Into Its Mechanism(s) of ActionwhNoch keine Bewertungen
- MenopauseDokument21 SeitenMenopauseDr K AmbareeshaNoch keine Bewertungen
- Changes in Leptin in Relation To Increased Testosterone Levels Associated With Eurycoma Longifolia Jack (Tongkat..Dokument4 SeitenChanges in Leptin in Relation To Increased Testosterone Levels Associated With Eurycoma Longifolia Jack (Tongkat..Hitesh VermaNoch keine Bewertungen
- Effect of Eurycoma Longifolia Standardised Aqueous Root Extract - Physta® On Testosterone Levels and Quality of LifeDokument15 SeitenEffect of Eurycoma Longifolia Standardised Aqueous Root Extract - Physta® On Testosterone Levels and Quality of LifeHavard MorkhagenNoch keine Bewertungen
- Eli GardDokument7 SeitenEli GardthanaNoch keine Bewertungen
- Body Reserves Affect The Reproductive enDokument13 SeitenBody Reserves Affect The Reproductive ensmokeridgeyvonneNoch keine Bewertungen
- tmp9099 TMPDokument8 Seitentmp9099 TMPFrontiersNoch keine Bewertungen
- Ashwagandha Hormonas SexualesDokument6 SeitenAshwagandha Hormonas SexualescumbredinNoch keine Bewertungen
- NIH Public Access: Author ManuscriptDokument20 SeitenNIH Public Access: Author ManuscriptDEShifNoch keine Bewertungen
- OligomenorheaDokument6 SeitenOligomenorhealianaNoch keine Bewertungen
- tmp9826 TMPDokument8 Seitentmp9826 TMPFrontiersNoch keine Bewertungen
- Glucocorticoids and Behavioral Effects of Psychostimulants. I: Locomotor Response To Cocaine Depends On Basal Levels of GlucocorticoidsDokument9 SeitenGlucocorticoids and Behavioral Effects of Psychostimulants. I: Locomotor Response To Cocaine Depends On Basal Levels of GlucocorticoidsSour60Noch keine Bewertungen
- KNOLL Enhancer Monograph 9 Nine 10 J 11 Ab J Bibl 1Dokument10 SeitenKNOLL Enhancer Monograph 9 Nine 10 J 11 Ab J Bibl 1JosephNoch keine Bewertungen
- 4 Karthick Et AlDokument4 Seiten4 Karthick Et AleditorijmrhsNoch keine Bewertungen
- 2562 - PDF OmigatDokument11 Seiten2562 - PDF OmigatkhansarafidaNoch keine Bewertungen
- Circulating Androgens in Women: Exercise-Induced ChangesDokument8 SeitenCirculating Androgens in Women: Exercise-Induced ChangesMuhamad Nazrul BoyoteenNoch keine Bewertungen
- Afro PilDokument14 SeitenAfro PilArchana SharmaNoch keine Bewertungen
- MelaDokument2 SeitenMelaChilly0104Noch keine Bewertungen
- The Long-Term Effects of Metoclopramide-Induced HyperprolactinemiaDokument7 SeitenThe Long-Term Effects of Metoclopramide-Induced Hyperprolactinemiaapi-282416840Noch keine Bewertungen
- 9 Ashutosh RanjanDokument10 Seiten9 Ashutosh RanjanAshutosh RanjanNoch keine Bewertungen
- Are Changes in MAPK ERK Necessary or Sufficient For Entrainment in Chick Pineal CellsDokument11 SeitenAre Changes in MAPK ERK Necessary or Sufficient For Entrainment in Chick Pineal CellsJose GarciaNoch keine Bewertungen
- Lorcaserin Summary by 4th GroupDokument3 SeitenLorcaserin Summary by 4th GroupdimbrutNoch keine Bewertungen
- Exam 19 Endocrine SystemDokument6 SeitenExam 19 Endocrine SystemPurwa RaneNoch keine Bewertungen
- Prof. Flavia Franconi: University of SassariDokument17 SeitenProf. Flavia Franconi: University of SassariSex & Gender Women's Health CollaborativeNoch keine Bewertungen
- Sibolone LBC Reference-3,5,6&7Dokument5 SeitenSibolone LBC Reference-3,5,6&7Kinjal ShahNoch keine Bewertungen
- Drug and Alcohol Dependence: Richard W. Foltin, Suzette M. EvansDokument10 SeitenDrug and Alcohol Dependence: Richard W. Foltin, Suzette M. EvansPaoChRNoch keine Bewertungen
- Medicinal Chemistry II Sovan Sarkar 186012111012Dokument11 SeitenMedicinal Chemistry II Sovan Sarkar 186012111012Sovan SarkarNoch keine Bewertungen
- Xenobiotic Regulation of Estrogen and Progesterone Receptor - Mediated Gene ExpressionVon EverandXenobiotic Regulation of Estrogen and Progesterone Receptor - Mediated Gene ExpressionNoch keine Bewertungen
- Diagnosis of Heavy Menstrual BleedingDokument6 SeitenDiagnosis of Heavy Menstrual BleedingLactose ScreamerNoch keine Bewertungen
- Norethindrone For The Jewish Bride: Grajower MSN COMDokument8 SeitenNorethindrone For The Jewish Bride: Grajower MSN COMoutdash2Noch keine Bewertungen
- Determinants of First Antenatal Visit Timing Among Pregnant Women in Antenatal Care at Kampala International University Teaching Hospital, Western UgandaDokument19 SeitenDeterminants of First Antenatal Visit Timing Among Pregnant Women in Antenatal Care at Kampala International University Teaching Hospital, Western UgandaKIU PUBLICATION AND EXTENSIONNoch keine Bewertungen
- Abnormalities-Of-Placenta-And-Umbilical CordDokument53 SeitenAbnormalities-Of-Placenta-And-Umbilical CordPraveen YadavNoch keine Bewertungen
- Genital ProlapseDokument39 SeitenGenital ProlapseFenias BoaneNoch keine Bewertungen
- Birth Control Comparison Chart 2018Dokument1 SeiteBirth Control Comparison Chart 2018Eric SandesNoch keine Bewertungen
- Birth PlanDokument3 SeitenBirth PlanMichaela ChamplinNoch keine Bewertungen
- Hydatidiform Mole: Incidence and Management Outcomes in A Tertiary Hospital in AbujaDokument5 SeitenHydatidiform Mole: Incidence and Management Outcomes in A Tertiary Hospital in AbujaSasa MicinNoch keine Bewertungen
- Obstetrics Gynaecology ReportDokument2 SeitenObstetrics Gynaecology ReportBEYOND ZFSNoch keine Bewertungen
- Teenage PregnancyDokument3 SeitenTeenage Pregnancyliezel napoles100% (1)
- Population Explosion-: Family Welfare ProgrammmeDokument15 SeitenPopulation Explosion-: Family Welfare ProgrammmeAiyanNoch keine Bewertungen
- Female Reproductive Anatomy: Unit 3: Female Reproducti Ve System 1Dokument33 SeitenFemale Reproductive Anatomy: Unit 3: Female Reproducti Ve System 1ERIC ASAMAMLEH KLEMEHNoch keine Bewertungen
- Faktor Yang Mempengaruhi Rendahnya Penggunaan KB Implant Pada Wanita Pasangan Usia Subur Di Puskemas BelawanDokument8 SeitenFaktor Yang Mempengaruhi Rendahnya Penggunaan KB Implant Pada Wanita Pasangan Usia Subur Di Puskemas Belawansarifah wildaNoch keine Bewertungen
- Holly TranzorDokument2 SeitenHolly Tranzorapi-26246128Noch keine Bewertungen
- Making Pregnancy Safer Builds On The Safe MotherhoodDokument17 SeitenMaking Pregnancy Safer Builds On The Safe MotherhoodRahmad ParadeNoch keine Bewertungen
- Research FINAL-wordDokument23 SeitenResearch FINAL-wordJahnenNoch keine Bewertungen
- What We Know About Primary Dysmenorrhea Today - A Critical ReviewDokument17 SeitenWhat We Know About Primary Dysmenorrhea Today - A Critical ReviewEva Aviles AlvarezNoch keine Bewertungen
- DR - Novan-Preterm Birth Awam 2Dokument31 SeitenDR - Novan-Preterm Birth Awam 2Mamik ShofiaNoch keine Bewertungen
- Chronic Pelvic Pain Women's Health Case ReportDokument6 SeitenChronic Pelvic Pain Women's Health Case ReportSharon Yunn Li ChenNoch keine Bewertungen
- Artéria Umbilical - IP Acharya 2005Dokument1 SeiteArtéria Umbilical - IP Acharya 2005Ruy Daniel Ferreira SouzaNoch keine Bewertungen
- RPM ACOG Practice BulletinDokument13 SeitenRPM ACOG Practice BulletinGabriela Cordova GoicocheaNoch keine Bewertungen
- 2322UN Women Analysis On Women and SDGsDokument32 Seiten2322UN Women Analysis On Women and SDGsNisaNoch keine Bewertungen