Das könnte Ihnen auch gefallen
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (890)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (587)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (265)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2219)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (119)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- A Design and Analysis of A Morphing Hyper-Elliptic Cambered Span (HECS) WingDokument10 SeitenA Design and Analysis of A Morphing Hyper-Elliptic Cambered Span (HECS) WingJEORJENoch keine Bewertungen
- 12 Animation Q1 AnswerDokument38 Seiten12 Animation Q1 AnswerBilly Joe TanNoch keine Bewertungen
- Instrumentation Design UTHMDokument5 SeitenInstrumentation Design UTHMAnis AzwaNoch keine Bewertungen
- Control PhilosophyDokument2 SeitenControl PhilosophytsplinstNoch keine Bewertungen
- LC IN SCIENCE and MathDokument14 SeitenLC IN SCIENCE and MathCharity Anne Camille PenalozaNoch keine Bewertungen
- Mitsubishi diesel forklifts 1.5-3.5 tonnesDokument2 SeitenMitsubishi diesel forklifts 1.5-3.5 tonnesJoniNoch keine Bewertungen
- Onishi T Ed Quantum Science The Frontier of Physics and ChemDokument498 SeitenOnishi T Ed Quantum Science The Frontier of Physics and ChemStrahinja DonicNoch keine Bewertungen
- Dollar Bill David Star - Verbal DiagramDokument4 SeitenDollar Bill David Star - Verbal DiagramLuciffer ChongNoch keine Bewertungen
- Physical parameters shaping of Farwa Lagoon - LibyaDokument7 SeitenPhysical parameters shaping of Farwa Lagoon - LibyaAsadeg ZaidNoch keine Bewertungen
- Reference Mil-Aero Guide ConnectorDokument80 SeitenReference Mil-Aero Guide ConnectorjamesclhNoch keine Bewertungen
- AMD OverDrive Utility 4.3.1.0698 Release NotesDokument1 SeiteAMD OverDrive Utility 4.3.1.0698 Release Notesbayu_gendeng666Noch keine Bewertungen
- Methods For Assessing The Stability of Slopes During Earthquakes-A Retrospective 1Dokument3 SeitenMethods For Assessing The Stability of Slopes During Earthquakes-A Retrospective 1ilijarskNoch keine Bewertungen
- BetaDokument16 SeitenBetaAkshita Saxena100% (2)
- Pragathi Full Test-6, Paper-2 (Qp+Key) - 18.07.22Dokument14 SeitenPragathi Full Test-6, Paper-2 (Qp+Key) - 18.07.22Ganesan MuthukrishnanNoch keine Bewertungen
- Scramjet EngineDokument2 SeitenScramjet EngineSãröj ShâhNoch keine Bewertungen
- Superconductivity in RH S and PD Se: A Comparative StudyDokument5 SeitenSuperconductivity in RH S and PD Se: A Comparative StudyChithra ArulmozhiNoch keine Bewertungen
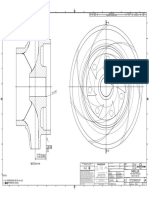
- Impeller: REV Rev by Description PCN / Ecn Date CHK'D A JMM Released For Production N/A 18/11/2019 PDLDokument1 SeiteImpeller: REV Rev by Description PCN / Ecn Date CHK'D A JMM Released For Production N/A 18/11/2019 PDLSenthilkumar RamalingamNoch keine Bewertungen
- Haidarali-MR-2011-PhD-Thesis 01 PDFDokument378 SeitenHaidarali-MR-2011-PhD-Thesis 01 PDFIbrahim KhanNoch keine Bewertungen
- Geotehnical Engg. - AEE - CRPQsDokument48 SeitenGeotehnical Engg. - AEE - CRPQsSureshKonamNoch keine Bewertungen
- San Unit-Wise QuestionsDokument6 SeitenSan Unit-Wise QuestionsSushmitha KodupakaNoch keine Bewertungen
- Drager Fabius Gs Technical Service ManualDokument350 SeitenDrager Fabius Gs Technical Service ManualLeonardo Chirinos100% (3)
- Training Matrix For TM IDokument14 SeitenTraining Matrix For TM IApril NavaretteNoch keine Bewertungen
- Control Lights with SOU-1 Twilight SwitchDokument1 SeiteControl Lights with SOU-1 Twilight SwitchjsblNoch keine Bewertungen
- Astm A6 A6m-08Dokument62 SeitenAstm A6 A6m-08Vũ Nhân HòaNoch keine Bewertungen
- Analisis Pengaruh Profitabilitas, Strategi Diversifikasi, Dan Good Corporate Governance Terhadap Nilai PerusahaanDokument16 SeitenAnalisis Pengaruh Profitabilitas, Strategi Diversifikasi, Dan Good Corporate Governance Terhadap Nilai PerusahaanEra ZsannabelaNoch keine Bewertungen
- © Ncert Not To Be Republished: AlgebraDokument12 Seiten© Ncert Not To Be Republished: Algebrakritagyasharma29Noch keine Bewertungen
- Visualizing Interstellar WormholesDokument14 SeitenVisualizing Interstellar WormholesFranciscoNoch keine Bewertungen
- WR424GB00DDokument16 SeitenWR424GB00DIgor San Martín PeñalozaNoch keine Bewertungen
- Bellin, E. H. (1984) - The Psychoanalytic Narrative On The Transformational Axis Between Writing and SpeechDokument15 SeitenBellin, E. H. (1984) - The Psychoanalytic Narrative On The Transformational Axis Between Writing and SpeechofanimenochNoch keine Bewertungen
- Sec 2 French ImmersionDokument1 SeiteSec 2 French Immersionapi-506328259Noch keine Bewertungen