Das könnte Ihnen auch gefallen
- Orthopaedics and Trauma for Medical Students and Junior ResidentsVon EverandOrthopaedics and Trauma for Medical Students and Junior ResidentsBewertung: 5 von 5 Sternen5/5 (3)
- Bone Development and Growth: Rosy Setiawati and Paulus RahardjoDokument20 SeitenBone Development and Growth: Rosy Setiawati and Paulus Rahardjoputri aisheNoch keine Bewertungen
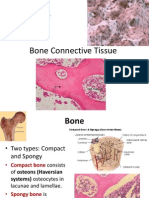
- Bones As A Living Dynamic TissueDokument13 SeitenBones As A Living Dynamic TissueSanish Basnet100% (1)
- BONE (Histology)Dokument65 SeitenBONE (Histology)fhfebriiNoch keine Bewertungen
- Alv - Bone FinalDokument28 SeitenAlv - Bone Finalv.shivakumarNoch keine Bewertungen
- Foc 2003 14 2 9Dokument8 SeitenFoc 2003 14 2 9Andy HongNoch keine Bewertungen
- Basic Bone BiologyDokument3 SeitenBasic Bone BiologyRuxandra Maria100% (1)
- FRACTURESDokument85 SeitenFRACTURESSteven OdhiamboNoch keine Bewertungen
- Fracture HealingDokument40 SeitenFracture Healingmohammad farhanNoch keine Bewertungen
- Dr. Sassia Lecture Bone Histology Part II July 4 2023Dokument39 SeitenDr. Sassia Lecture Bone Histology Part II July 4 2023Ali ELKARGHALYNoch keine Bewertungen
- Normal Bone Anatomy and Physiology: Go ToDokument11 SeitenNormal Bone Anatomy and Physiology: Go TotariNoch keine Bewertungen
- Cell Lines and Primary Cell Cultures in The Study of Bone Cell BiologyDokument24 SeitenCell Lines and Primary Cell Cultures in The Study of Bone Cell BiologyAngelNoch keine Bewertungen
- DO - DPM Bone Formation D2L 2014Dokument7 SeitenDO - DPM Bone Formation D2L 2014snlee06Noch keine Bewertungen
- Osteoblasts, Osteoclasts, and Osteocytes: Unveiling Their Intimate-Associated Responses To Applied Orthodontic ForcesDokument12 SeitenOsteoblasts, Osteoclasts, and Osteocytes: Unveiling Their Intimate-Associated Responses To Applied Orthodontic ForcesMariya NazimNoch keine Bewertungen
- Histogenesis of BoneDokument6 SeitenHistogenesis of BoneAlya Putri KhairaniNoch keine Bewertungen
- Ch6Bone StudentDokument66 SeitenCh6Bone Studentbhattiareeba13Noch keine Bewertungen
- Osteogenesis: The Development of Bones: Intramembranous OssificationDokument9 SeitenOsteogenesis: The Development of Bones: Intramembranous OssificationihsansiregarNoch keine Bewertungen
- 2) Development and Growth of The BonesDokument3 Seiten2) Development and Growth of The BonesAris PaparisNoch keine Bewertungen
- Bone and Cartilage TissueDokument33 SeitenBone and Cartilage TissueGIANG ĐẶNG DUY TRÚCNoch keine Bewertungen
- HHS Public Access: The Potential of Probiotics As A Therapy For OsteoporosisDokument26 SeitenHHS Public Access: The Potential of Probiotics As A Therapy For OsteoporosisBagastyoNoch keine Bewertungen
- Ch6Bone StudentDokument67 SeitenCh6Bone Studentbhattiareeba13Noch keine Bewertungen
- Ossification: Ossification (Or Osteogenesis) in Bone Remodeling Is TheDokument3 SeitenOssification: Ossification (Or Osteogenesis) in Bone Remodeling Is TheAlan FosterNoch keine Bewertungen
- Biochemistry of Bone (Assignment)Dokument7 SeitenBiochemistry of Bone (Assignment)haseeb ShafaatNoch keine Bewertungen
- TARI ReferensiDokument13 SeitenTARI ReferensitariNoch keine Bewertungen
- Chapter 1Dokument21 SeitenChapter 1Fernando Melo GarciaNoch keine Bewertungen
- Fracture Healing and BonegraftingDokument76 SeitenFracture Healing and BonegraftingJio AmurNoch keine Bewertungen
- Bone Structure: OsteoblastsDokument4 SeitenBone Structure: OsteoblastsamlniNoch keine Bewertungen
- Bone Anatomy Good NotesDokument16 SeitenBone Anatomy Good NotesDENACE JnrNoch keine Bewertungen
- Bone FormationDokument9 SeitenBone FormationKamalakannanNoch keine Bewertungen
- Bone Development: Osteogenesis (Ossification) Endochondral OssificationDokument2 SeitenBone Development: Osteogenesis (Ossification) Endochondral OssificationVanshika SethiNoch keine Bewertungen
- Bones Ossification.Dokument9 SeitenBones Ossification.Shimmering MoonNoch keine Bewertungen
- Normal Bone Anatomy and PhysiologyDokument9 SeitenNormal Bone Anatomy and PhysiologyElsa GonçalvesNoch keine Bewertungen
- Bones OssificationDokument9 SeitenBones OssificationNaveed AfridiNoch keine Bewertungen
- BONEDokument48 SeitenBONEnivethaseshaNoch keine Bewertungen
- 05 Locomotor SkeletalDokument288 Seiten05 Locomotor SkeletalkalumbaaquilaNoch keine Bewertungen
- BONE - Copy 2Dokument27 SeitenBONE - Copy 2negmm2226Noch keine Bewertungen
- Ossification of Bone.Dokument22 SeitenOssification of Bone.Shimmering MoonNoch keine Bewertungen
- Osteoblast: osteo-and βλαστάνω, blastanō "germinate") are cells with a singleDokument10 SeitenOsteoblast: osteo-and βλαστάνω, blastanō "germinate") are cells with a singleAlan FosterNoch keine Bewertungen
- Pathologic Bone FractureDokument5 SeitenPathologic Bone FractureJaysellePuguonTabijeNoch keine Bewertungen
- Skeletal SystemDokument7 SeitenSkeletal SystemFlorilyn CabezoNoch keine Bewertungen
- Skeletal System More On ProcessDokument4 SeitenSkeletal System More On Processsayie potatoNoch keine Bewertungen
- Fracture Healing in Cortical and Cancellous BoneDokument63 SeitenFracture Healing in Cortical and Cancellous BonePramod Thottimane0% (1)
- Bone Growth and Factors That Associated With It1234Dokument7 SeitenBone Growth and Factors That Associated With It1234Amanuel TarekegnNoch keine Bewertungen
- Principles of Bone GraftingDokument6 SeitenPrinciples of Bone GraftingJayanth Perumal100% (2)
- Bone Healing ProcessDokument21 SeitenBone Healing ProcessWing YenNoch keine Bewertungen
- The Osteocyte: From "Prisoner" To "Orchestrator": Carla Palumbo and Marzia FerrettiDokument17 SeitenThe Osteocyte: From "Prisoner" To "Orchestrator": Carla Palumbo and Marzia FerrettiNadira NurinNoch keine Bewertungen
- Biomechanics and Biomaterials in Orthopedic SurgeryDokument11 SeitenBiomechanics and Biomaterials in Orthopedic SurgeryManiventhan NachimuthuNoch keine Bewertungen
- 09lect Bone Summary2017Dokument3 Seiten09lect Bone Summary2017Dan ChoiNoch keine Bewertungen
- Nihms-1641529 Mechanisms of Bone Development and RepairDokument41 SeitenNihms-1641529 Mechanisms of Bone Development and Repairhennysusanto18Noch keine Bewertungen
- Practical Work No. 7: Faculty of MedicineDokument3 SeitenPractical Work No. 7: Faculty of MedicineAndrei TimofteNoch keine Bewertungen
- LP 7 Connective Tissue BoneDokument3 SeitenLP 7 Connective Tissue BoneAndrei TimofteNoch keine Bewertungen
- Bone Tissue-Growth and DevelopmentDokument53 SeitenBone Tissue-Growth and Developmentgalaxyduck100% (1)
- 11 Residual Ridge ResorptionDokument66 Seiten11 Residual Ridge ResorptionSuman JainNoch keine Bewertungen
- Arnett, T.Dokument10 SeitenArnett, T.Alex AmorimNoch keine Bewertungen
- PIIS0030589809000881Dokument12 SeitenPIIS0030589809000881dr. Joko WibowoNoch keine Bewertungen
- BIOL3307 - Unit1Mod2 - Bone Growth & Repair1Dokument16 SeitenBIOL3307 - Unit1Mod2 - Bone Growth & Repair1Ren StudyNoch keine Bewertungen
- Skeletal System Lesson 2Dokument40 SeitenSkeletal System Lesson 2Ella Nika FangonNoch keine Bewertungen
- OssificationDokument3 SeitenOssificationFatin Nabihah JamilNoch keine Bewertungen
- 7 - BoneDokument36 Seiten7 - BoneKhdhir M. SalemNoch keine Bewertungen
- Draft Manuscript Uncaria Gambir Tanpa Diskusi Edit SalsaDokument20 SeitenDraft Manuscript Uncaria Gambir Tanpa Diskusi Edit SalsaSalsabila AfniaNoch keine Bewertungen
- Catalogue: Oem/OdmDokument14 SeitenCatalogue: Oem/OdmCarlito RiccyNoch keine Bewertungen
- Orthopedics BoneDisordersDokument20 SeitenOrthopedics BoneDisordersannapanna1Noch keine Bewertungen
- Biomechanics of Human Bone Growth & DevelopmentDokument40 SeitenBiomechanics of Human Bone Growth & DevelopmentShimmering MoonNoch keine Bewertungen
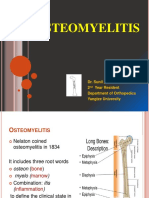
- Osteomyelitis: Dr. Sunil Pahari 2 Year Resident Department of Orthopedics Yangtze UniversityDokument48 SeitenOsteomyelitis: Dr. Sunil Pahari 2 Year Resident Department of Orthopedics Yangtze UniversityPercy Linares MorilloNoch keine Bewertungen
- Anatomy Yoga Therapy Conf ManualDokument84 SeitenAnatomy Yoga Therapy Conf ManualPablo ErpelNoch keine Bewertungen
- Pregnancy & OrthodonticsDokument5 SeitenPregnancy & OrthodonticsAnonymous Y9FLG5kQdNoch keine Bewertungen
- Hip DislocationDokument39 SeitenHip DislocationJordan Garcia Aguilar0% (1)
- ANPH-M1-CU4. Skeletal SystemDokument26 SeitenANPH-M1-CU4. Skeletal SystemajdNoch keine Bewertungen
- Skeletal System PDFDokument24 SeitenSkeletal System PDFJennifer LalondeNoch keine Bewertungen
- Cell SaltsDokument8 SeitenCell SaltsminunatNoch keine Bewertungen
- Impact of Radiotherapy On Mandibular Bone: A Retrospective Study of Digital Panoramic RadiographsDokument6 SeitenImpact of Radiotherapy On Mandibular Bone: A Retrospective Study of Digital Panoramic Radiographsboooow92Noch keine Bewertungen
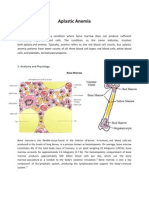
- Aplastic AnemiaDokument5 SeitenAplastic AnemiaVenice Marie GargantaNoch keine Bewertungen
- Brief History of Osteoarthritis and Back PainDokument2 SeitenBrief History of Osteoarthritis and Back PainAnnaNoch keine Bewertungen
- Musculoskeletal QuizDokument4 SeitenMusculoskeletal Quizpapa.pradoNoch keine Bewertungen
- C. Andrew L. Bassett - Beneficial Effects of Electromagnetic FieldsDokument7 SeitenC. Andrew L. Bassett - Beneficial Effects of Electromagnetic FieldsDsgmmmmNoch keine Bewertungen
- MA Bone CellsDokument10 SeitenMA Bone CellskristinaNoch keine Bewertungen
- Skeleton. A&PDokument3 SeitenSkeleton. A&Pbayoagunbiade0Noch keine Bewertungen
- Medical Surgical Nursing Orthopedic NursingDokument141 SeitenMedical Surgical Nursing Orthopedic Nursingroger92% (12)
- RT 112 Module 2Dokument67 SeitenRT 112 Module 2AVE MAE ALEXIS NAVATANoch keine Bewertungen
- Metabolic Bone ConditionsDokument66 SeitenMetabolic Bone ConditionsNaeem AminNoch keine Bewertungen
- 4 AnaDokument22 Seiten4 Anairis casalaNoch keine Bewertungen
- Healing of FracturesDokument19 SeitenHealing of FracturesTracy100% (4)
- The Importance of Friendship EssayDokument6 SeitenThe Importance of Friendship Essayafibojmbjifexj100% (2)
- MODULE 4 Skeletal SystemDokument16 SeitenMODULE 4 Skeletal SystemKate Andrea PanizalesNoch keine Bewertungen
- Syllabus Basic B.SCDokument232 SeitenSyllabus Basic B.SCkiran kumarNoch keine Bewertungen
- ANAPHY Lec Session #3 - SASDokument12 SeitenANAPHY Lec Session #3 - SASFherry Mae UsmanNoch keine Bewertungen
- 14 - Mazouz - Cell Free Qantitative Mineralization Measurements To Identify OI BGSDokument12 Seiten14 - Mazouz - Cell Free Qantitative Mineralization Measurements To Identify OI BGSfarida.harouniiNoch keine Bewertungen
- Large Fragment Locking Compression Plate (LCP) : Technique GuideDokument23 SeitenLarge Fragment Locking Compression Plate (LCP) : Technique GuideLouis MiuNoch keine Bewertungen
- Manifestaciones Reumatológicas de Enfermedades EndocrinológicasDokument10 SeitenManifestaciones Reumatológicas de Enfermedades EndocrinológicasMarcela Catalina Fandiño VargasNoch keine Bewertungen
- Love Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)Von EverandLove Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)Bewertung: 3 von 5 Sternen3/5 (1)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedVon EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedBewertung: 4.5 von 5 Sternen4.5/5 (82)
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDVon EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDBewertung: 5 von 5 Sternen5/5 (3)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionVon EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionBewertung: 4 von 5 Sternen4/5 (404)
- The Age of Magical Overthinking: Notes on Modern IrrationalityVon EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityBewertung: 4 von 5 Sternen4/5 (32)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeVon EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeBewertung: 2 von 5 Sternen2/5 (1)
- Manipulation: The Ultimate Guide To Influence People with Persuasion, Mind Control and NLP With Highly Effective Manipulation TechniquesVon EverandManipulation: The Ultimate Guide To Influence People with Persuasion, Mind Control and NLP With Highly Effective Manipulation TechniquesBewertung: 4.5 von 5 Sternen4.5/5 (1412)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsVon EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNoch keine Bewertungen
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsVon EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsBewertung: 4 von 5 Sternen4/5 (4)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsVon EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsBewertung: 5 von 5 Sternen5/5 (1)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisVon EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisBewertung: 4.5 von 5 Sternen4.5/5 (42)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaVon EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Comfort of Crows: A Backyard YearVon EverandThe Comfort of Crows: A Backyard YearBewertung: 4.5 von 5 Sternen4.5/5 (23)
- The Obesity Code: Unlocking the Secrets of Weight LossVon EverandThe Obesity Code: Unlocking the Secrets of Weight LossBewertung: 4 von 5 Sternen4/5 (6)
- Why We Die: The New Science of Aging and the Quest for ImmortalityVon EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityBewertung: 4.5 von 5 Sternen4.5/5 (6)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisVon EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisBewertung: 3.5 von 5 Sternen3.5/5 (2)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeVon EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeBewertung: 4.5 von 5 Sternen4.5/5 (254)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Von EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Bewertung: 4.5 von 5 Sternen4.5/5 (110)
- To Explain the World: The Discovery of Modern ScienceVon EverandTo Explain the World: The Discovery of Modern ScienceBewertung: 3.5 von 5 Sternen3.5/5 (51)
- Critical Thinking: How to Effectively Reason, Understand Irrationality, and Make Better DecisionsVon EverandCritical Thinking: How to Effectively Reason, Understand Irrationality, and Make Better DecisionsBewertung: 4.5 von 5 Sternen4.5/5 (39)
- The Marshmallow Test: Mastering Self-ControlVon EverandThe Marshmallow Test: Mastering Self-ControlBewertung: 4.5 von 5 Sternen4.5/5 (60)
- Dark Psychology: Learn To Influence Anyone Using Mind Control, Manipulation And Deception With Secret Techniques Of Dark Persuasion, Undetected Mind Control, Mind Games, Hypnotism And BrainwashingVon EverandDark Psychology: Learn To Influence Anyone Using Mind Control, Manipulation And Deception With Secret Techniques Of Dark Persuasion, Undetected Mind Control, Mind Games, Hypnotism And BrainwashingBewertung: 4 von 5 Sternen4/5 (1138)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessVon EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessBewertung: 4.5 von 5 Sternen4.5/5 (328)