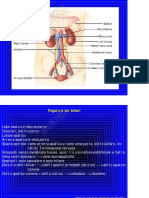
Das könnte Ihnen auch gefallen
- Gas Tri TiDokument57 SeitenGas Tri Tiapi-3733154100% (1)
- Lezione Ormoni GastrointestinaliDokument28 SeitenLezione Ormoni GastrointestinaliAnna De SimoneNoch keine Bewertungen
- Digestione Monogastrici RuminantiDokument54 SeitenDigestione Monogastrici RuminantiEnea VirgilioNoch keine Bewertungen
- 3.digestione Monogastrici Ruminanti2014Dokument60 Seiten3.digestione Monogastrici Ruminanti2014Alessandra Patricia LopesNoch keine Bewertungen
- Farmaco 1Dokument15 SeitenFarmaco 1AlessandraManuela VisentinNoch keine Bewertungen
- Fisiologia e Fisiopatologia Del Diabete MellitoDokument10 SeitenFisiologia e Fisiopatologia Del Diabete Mellitosiluma11100% (1)
- Sbobina Lezione 15 BiochimicaDokument15 SeitenSbobina Lezione 15 BiochimicaGiorgia de SantisNoch keine Bewertungen
- Uomo e NaturaDokument6 SeitenUomo e NaturaAlessia PetragalloNoch keine Bewertungen
- Medicina InternaDokument112 SeitenMedicina InternaAutogeneratasiNoch keine Bewertungen
- Ormoni Gastrointestinali 2Dokument25 SeitenOrmoni Gastrointestinali 2Anna De SimoneNoch keine Bewertungen
- Amminoacidi e ProteineDokument64 SeitenAmminoacidi e ProteineMajorana.WEBNoch keine Bewertungen
- LEZIONE 9 Ossidazione Acidi GrassiDokument58 SeitenLEZIONE 9 Ossidazione Acidi GrassiMaegazine EnizageamNoch keine Bewertungen
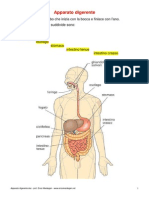
- Apparato DigerenteDokument10 SeitenApparato DigerenteBruce WiiNoch keine Bewertungen
- Metabolismo Delle ProteineDokument10 SeitenMetabolismo Delle ProteineGianluca Garozzo100% (1)
- Omeopatia - Lo Stomaco E Le Piante MedicinaliDokument91 SeitenOmeopatia - Lo Stomaco E Le Piante MedicinaliLuca MauraNoch keine Bewertungen
- Apparato DigerenteDokument11 SeitenApparato DigerenteMary CotugnoNoch keine Bewertungen
- Proteine AlDokument66 SeitenProteine Alrinodelgattopardolibero.itNoch keine Bewertungen
- Catabolismo AminoacidiDokument6 SeitenCatabolismo AminoacidiDavide BianchiNoch keine Bewertungen
- Med_Odonto_METABOLISMO_DELLE_BASI_PURINICHE_E_PIRIMIDINICHEDokument93 SeitenMed_Odonto_METABOLISMO_DELLE_BASI_PURINICHE_E_PIRIMIDINICHEFilippo BartoliNoch keine Bewertungen
- Coenzimi e VitamineDokument37 SeitenCoenzimi e VitamineRoberta Lo IzzoNoch keine Bewertungen
- Riassunto VitamineDokument7 SeitenRiassunto VitamineStefano0% (1)
- Lezione 9Dokument54 SeitenLezione 9Tonia PreziusoNoch keine Bewertungen
- Apparato DigerenteDokument7 SeitenApparato DigerenteCatalina Valeria MocanuNoch keine Bewertungen
- 01a Cenni Di Anatomia e Fisiologia Dell'Apparato DigerenteDokument5 Seiten01a Cenni Di Anatomia e Fisiologia Dell'Apparato DigerenteFrancesco NaniaNoch keine Bewertungen
- GottaDokument9 SeitenGottaannaricci.aNoch keine Bewertungen
- 12 Gastroenterologia PANCREATITIDokument13 Seiten12 Gastroenterologia PANCREATITIangelmartinez87Noch keine Bewertungen
- Metabolismo Calcio FosforoDokument6 SeitenMetabolismo Calcio Fosforothe_answer911572Noch keine Bewertungen
- Combinazioni Alimentari PDFDokument5 SeitenCombinazioni Alimentari PDFmarco poloNoch keine Bewertungen
- Biochimica Del FegatoDokument6 SeitenBiochimica Del FegatoSalò MarzialiNoch keine Bewertungen
- Patologia Clinica FegatoDokument14 SeitenPatologia Clinica FegatoBarbara BranconeNoch keine Bewertungen
- A EnzimiDokument15 SeitenA EnzimiBiagio CastronovoNoch keine Bewertungen
- DigestioneDokument11 SeitenDigestionesono bruttoNoch keine Bewertungen
- Sistema Endocrino e Ormoni Prof. Marco GiammancoDokument59 SeitenSistema Endocrino e Ormoni Prof. Marco GiammancoMarco GiammancoNoch keine Bewertungen
- Biochimica AvanzataDokument15 SeitenBiochimica Avanzataapi-3863584100% (3)
- Apparato DigerenteDokument58 SeitenApparato DigerenteGiorgio100% (1)
- GastroenterologiaDokument12 SeitenGastroenterologianadiaNoch keine Bewertungen
- LimpidexDokument99 SeitenLimpidexmarioNoch keine Bewertungen
- Sbobina CURS 5Dokument5 SeitenSbobina CURS 5Giorgia ManieroNoch keine Bewertungen
- Controllo MetabolismoDokument31 SeitenControllo Metabolismosweet_05Noch keine Bewertungen
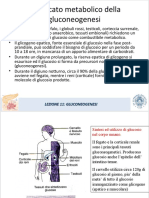
- LEZIONE 11 Gluconeogenesi SMODokument44 SeitenLEZIONE 11 Gluconeogenesi SMOMaegazine EnizageamNoch keine Bewertungen
- Elenco Quesiti Biochimica ClinicaDokument10 SeitenElenco Quesiti Biochimica ClinicaNicola LauroraNoch keine Bewertungen
- Vitamine IdrosolubileDokument30 SeitenVitamine IdrosolubileMarco GiNoch keine Bewertungen
- Sistema Gastrointestinale Fisiologia DelDokument160 SeitenSistema Gastrointestinale Fisiologia DelMiriam GalliNoch keine Bewertungen
- Apparato DigerenteDokument104 SeitenApparato Digerentesono bruttoNoch keine Bewertungen
- 5-Fibra AlimentareDokument27 Seiten5-Fibra Alimentarepappo pappaNoch keine Bewertungen
- Biochimica ClinicaDokument37 SeitenBiochimica ClinicaMarco OddeninoNoch keine Bewertungen
- 6 Lezione ScalfiDokument8 Seiten6 Lezione ScalfiClaudio MormoneNoch keine Bewertungen
- Tesi Roberta Musumeci 9 03 2012Dokument78 SeitenTesi Roberta Musumeci 9 03 2012api-322196723Noch keine Bewertungen
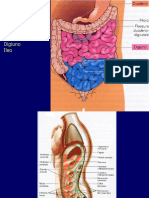
- INTESTINO TENUEDokument6 SeitenINTESTINO TENUEHghh NgthhNoch keine Bewertungen
- Disbiosi Intestinale e Patologie Correlate DR - Lozio LucianoDokument100 SeitenDisbiosi Intestinale e Patologie Correlate DR - Lozio LucianoCosimo Lanza100% (2)
- 20 Le Aferesi Selettive - TecnologiaDokument40 Seiten20 Le Aferesi Selettive - TecnologiaAndreaNoch keine Bewertungen
- DietologiaDokument16 SeitenDietologiamajholx0% (1)
- Lezione 23 Da StamapreDokument6 SeitenLezione 23 Da StamapreMariacarmela MorrealeNoch keine Bewertungen
- Integrazione Alimetare Nelle Allergie e Intolleranze Alimemtari - Integrazione FunzionaleDokument66 SeitenIntegrazione Alimetare Nelle Allergie e Intolleranze Alimemtari - Integrazione Funzionalemaria maggioNoch keine Bewertungen
- 2021 Farmacia Digerente 2 23 26 AprileDokument78 Seiten2021 Farmacia Digerente 2 23 26 AprileChiara ArdauNoch keine Bewertungen
- Gastroeneterologia Sunto FinalefDokument11 SeitenGastroeneterologia Sunto FinalefNunzio OlivieroNoch keine Bewertungen
- BIOCHIMICA - SiliprandiDokument50 SeitenBIOCHIMICA - SiliprandiinfernuxNoch keine Bewertungen
- Tumori SNCDokument8 SeitenTumori SNCNoemi Di FucciaNoch keine Bewertungen
- Principi Di EmodinamicaDokument17 SeitenPrincipi Di EmodinamicaNoemi Di FucciaNoch keine Bewertungen
- Lesioni Elementari Della CuteDokument9 SeitenLesioni Elementari Della CuteNoemi Di FucciaNoch keine Bewertungen
- Test Funzionalità RenaleDokument54 SeitenTest Funzionalità RenaleNoemi Di FucciaNoch keine Bewertungen
- Lezioni Fisiologia CardiacaDokument18 SeitenLezioni Fisiologia CardiacaNoemi Di FucciaNoch keine Bewertungen
- Domande FisiologiaDokument6 SeitenDomande FisiologiaNoemi Di FucciaNoch keine Bewertungen
- Controllo Della CircolazioneDokument26 SeitenControllo Della CircolazioneNoemi Di FucciaNoch keine Bewertungen
- Meccanica RespiratoriaDokument23 SeitenMeccanica RespiratoriaNoemi Di FucciaNoch keine Bewertungen
- Semeiotica PancreasDokument31 SeitenSemeiotica PancreasNoemi Di FucciaNoch keine Bewertungen
- Statistica MedicaDokument2 SeitenStatistica MedicaNoemi Di FucciaNoch keine Bewertungen
- Statistica Medica 15 Novembre 2012Dokument5 SeitenStatistica Medica 15 Novembre 2012Noemi Di FucciaNoch keine Bewertungen
- Microbiologia - Virologia 19 AprileDokument5 SeitenMicrobiologia - Virologia 19 AprileNoemi Di FucciaNoch keine Bewertungen
- Named Articolo Prof Dioguardi Indicazioni Uso Aminoacidi-Essenziali 2020Dokument4 SeitenNamed Articolo Prof Dioguardi Indicazioni Uso Aminoacidi-Essenziali 2020Vera SavastanoNoch keine Bewertungen
- Regolazione Dell'Espressione GenicaLICEODokument72 SeitenRegolazione Dell'Espressione GenicaLICEOReaperguyNoch keine Bewertungen
- Merceologia Produselor AlimentareDokument49 SeitenMerceologia Produselor AlimentareAttil TorokNoch keine Bewertungen
- (S7ita - Bi2ita) Presentazione: La ProteosintesiDokument26 Seiten(S7ita - Bi2ita) Presentazione: La ProteosintesiSalomon C LeijaNoch keine Bewertungen
- Ciclo Di KrebsDokument13 SeitenCiclo Di KrebsLapo MosettiNoch keine Bewertungen
- Biologia MolecolareDokument180 SeitenBiologia MolecolareLorenzo Maria CerviNoch keine Bewertungen
- BiomolecoleDokument7 SeitenBiomolecolealessioanginiNoch keine Bewertungen
- BiomolecoleDokument3 SeitenBiomolecoleabcdefs96Noch keine Bewertungen