Das könnte Ihnen auch gefallen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- Spawn Production TechnologyDokument8 SeitenSpawn Production TechnologythosampaNoch keine Bewertungen
- Fish Breeding BookDokument84 SeitenFish Breeding Booksarveshnema100% (1)
- IB Biology HL - Option C Past PapersDokument32 SeitenIB Biology HL - Option C Past PapersTheIBGeoKid80% (5)
- 04 - Brood Stock Collection, Transport and MaintenanceDokument4 Seiten04 - Brood Stock Collection, Transport and MaintenanceNilamdeen Mohamed ZamilNoch keine Bewertungen
- Philippine Fisheries Code provisions on unauthorized fishingDokument105 SeitenPhilippine Fisheries Code provisions on unauthorized fishingDee LM0% (1)
- The Big Discus Fish BookDokument17 SeitenThe Big Discus Fish BookAndres Felipe Tirano VasquezNoch keine Bewertungen
- Speedy InstanDokument8 SeitenSpeedy InstanarifulkaNoch keine Bewertungen
- Username 2Dokument2 SeitenUsername 2arifulkaNoch keine Bewertungen
- Username 2Dokument2 SeitenUsername 2arifulkaNoch keine Bewertungen
- General Arrangement Mini Tug BoatDokument1 SeiteGeneral Arrangement Mini Tug BoatnawanNoch keine Bewertungen
- Aquatic Toxicology Volume 151 Issue 2014 [Doi 10.1016_j.aquatox.2013.10.012] Weinberger, Joel; Klaper, Rebecca -- Environmental Concentrations of the Selective Serotonin Reuptake Inhibitor Fluoxetine Impact Specif_2Dokument7 SeitenAquatic Toxicology Volume 151 Issue 2014 [Doi 10.1016_j.aquatox.2013.10.012] Weinberger, Joel; Klaper, Rebecca -- Environmental Concentrations of the Selective Serotonin Reuptake Inhibitor Fluoxetine Impact Specif_2arifulkaNoch keine Bewertungen
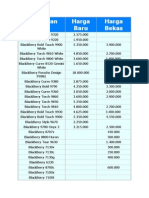
- Harga BBDokument5 SeitenHarga BBarifulkaNoch keine Bewertungen
- SpeedyDokument2 SeitenSpeedyarifulkaNoch keine Bewertungen
- No. Waktu Pengukuran (WIB) Ketinggian SatuanDokument2 SeitenNo. Waktu Pengukuran (WIB) Ketinggian SatuanarifulkaNoch keine Bewertungen
- Bhs Inggris TugasDokument4 SeitenBhs Inggris TugasarifulkaNoch keine Bewertungen
- No. Waktu Pengukuran (WIB) Ketinggian SatuanDokument2 SeitenNo. Waktu Pengukuran (WIB) Ketinggian SatuanarifulkaNoch keine Bewertungen
- Daftar Peserta SMPTN Mahasiswa Baru Pemanfaatan Sumerdaya Perikanan (111314) Tahun 2014Dokument2 SeitenDaftar Peserta SMPTN Mahasiswa Baru Pemanfaatan Sumerdaya Perikanan (111314) Tahun 2014arifulkaNoch keine Bewertungen
- Coral Reef Modelling DHI SolutionDokument2 SeitenCoral Reef Modelling DHI SolutionJonas BrandiNoch keine Bewertungen
- Induced spawning in fish: Effect of Ovaprim dosageDokument6 SeitenInduced spawning in fish: Effect of Ovaprim dosageMoaz FiazNoch keine Bewertungen
- Culture of Milkfish-TarpapelDokument7 SeitenCulture of Milkfish-TarpapelMaricel TandangNoch keine Bewertungen
- Aquaculture Potential of The Common OctopusDokument18 SeitenAquaculture Potential of The Common OctopusDoctorEmirNoch keine Bewertungen
- Oysters SsssDokument19 SeitenOysters SsssI'm a Smart CatNoch keine Bewertungen
- Contribution of Natural Plankton To The Diet of White Leg Shrimp Litopenaeus Vannamei (Boone, 1931) Post-Larvae in Fertilized Pond ConditionsDokument71 SeitenContribution of Natural Plankton To The Diet of White Leg Shrimp Litopenaeus Vannamei (Boone, 1931) Post-Larvae in Fertilized Pond ConditionsAlex ZomNoch keine Bewertungen
- RHM Baggani 112008Dokument4 SeitenRHM Baggani 112008Richard RossNoch keine Bewertungen
- Life in Lake Atitlan (Ati Divers, La Iguana Perdida, Guatemala)Dokument4 SeitenLife in Lake Atitlan (Ati Divers, La Iguana Perdida, Guatemala)Natalie Gillis100% (1)
- Sims 3 Create A World Instruction ManualDokument8 SeitenSims 3 Create A World Instruction ManualJuvy Rose Tinga YeeNoch keine Bewertungen
- FishDokument37 SeitenFishlambatiNoch keine Bewertungen
- A Manual For Commercial Production of The Gourami, Trichogaster Trichopterus - A Temporary Paired SpawnerDokument37 SeitenA Manual For Commercial Production of The Gourami, Trichogaster Trichopterus - A Temporary Paired SpawnernlribeiroNoch keine Bewertungen
- Biology and Life Cycle of Giant Tiger PrawnDokument13 SeitenBiology and Life Cycle of Giant Tiger PrawnAbir WahidNoch keine Bewertungen
- Danios and DevariosDokument2 SeitenDanios and DevariosREXTERYXNoch keine Bewertungen
- Egg Depositor of Ornamental FishesDokument4 SeitenEgg Depositor of Ornamental FishesPieter SteenkampNoch keine Bewertungen
- (Doi 10.1016 - b978-012374473-9.00026-6) Houde, E.D. - Encyclopedia of Ocean Sciences - Fish LarvaeDokument11 Seiten(Doi 10.1016 - b978-012374473-9.00026-6) Houde, E.D. - Encyclopedia of Ocean Sciences - Fish LarvaeAnkeet KumarNoch keine Bewertungen
- A Manual On Hatchery of Sea Cucumber Holothuria Scabra 3-LibreDokument21 SeitenA Manual On Hatchery of Sea Cucumber Holothuria Scabra 3-LibresuperaditrNoch keine Bewertungen
- Pathfinder 2e - Roll For Combat Indigo Isles Character Guide v1Dokument103 SeitenPathfinder 2e - Roll For Combat Indigo Isles Character Guide v1Michel Pinto de SousaNoch keine Bewertungen
- Broodstock and Hatchery Management: Ryan L. Lane, PHDDokument26 SeitenBroodstock and Hatchery Management: Ryan L. Lane, PHDUsman MadubunNoch keine Bewertungen
- Aquaculture Reports: A A B ADokument8 SeitenAquaculture Reports: A A B AThach Tuan AnhNoch keine Bewertungen
- Prathamesh 9 TH DDokument20 SeitenPrathamesh 9 TH Dnikita londhe100% (1)
- Oyster Farming in The PhilippinesDokument13 SeitenOyster Farming in The PhilippinesElizabeth B DiestroNoch keine Bewertungen
- Updates On Spawn Production of Volvariella Volvacea, The Tropical Straw MushroomDokument7 SeitenUpdates On Spawn Production of Volvariella Volvacea, The Tropical Straw MushroomAeroman MechanicNoch keine Bewertungen
- Coral Reef: Formation TypesDokument41 SeitenCoral Reef: Formation Typesenzo abrahamNoch keine Bewertungen
- Spat Collection StudyDokument8 SeitenSpat Collection StudynamdrikaleleNoch keine Bewertungen


















































![Aquatic Toxicology Volume 151 Issue 2014 [Doi 10.1016_j.aquatox.2013.10.012] Weinberger, Joel; Klaper, Rebecca -- Environmental Concentrations of the Selective Serotonin Reuptake Inhibitor Fluoxetine Impact Specif_2](https://imgv2-1-f.scribdassets.com/img/document/251151333/149x198/a3d1cbb703/1419742376?v=1)