Das könnte Ihnen auch gefallen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5795)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Jyotish - 1980 - Saha - Precious Stones That Heal PDFDokument258 SeitenJyotish - 1980 - Saha - Precious Stones That Heal PDFKrishan Sirohiya100% (1)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Circulatory System MCQDokument9 SeitenCirculatory System MCQFake GamerNoch keine Bewertungen
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- With Price DR LAL PATHLAB Final PDF - BookletDokument310 SeitenWith Price DR LAL PATHLAB Final PDF - BookletSurendra Singh Negi100% (1)
- Skin CancerDokument24 SeitenSkin CancerMister David anthonyNoch keine Bewertungen
- 1 - Hifumax 1Dokument25 Seiten1 - Hifumax 1Get It SuplementosNoch keine Bewertungen
- BPR in Posterior Tooth From A To ZDokument77 SeitenBPR in Posterior Tooth From A To ZPingyuan ChenNoch keine Bewertungen
- 0-1-How To Submit Aligner+ Consulting or Mentoring CasesDokument6 Seiten0-1-How To Submit Aligner+ Consulting or Mentoring CasesPingyuan ChenNoch keine Bewertungen
- Bleaching 1Dokument4 SeitenBleaching 1Pingyuan ChenNoch keine Bewertungen
- Can Intra-Coronally Bleached Teeth Be Bonded Safely After Antioxidant Treatment?Dokument6 SeitenCan Intra-Coronally Bleached Teeth Be Bonded Safely After Antioxidant Treatment?Pingyuan ChenNoch keine Bewertungen
- 767 FullDokument9 Seiten767 FullPingyuan ChenNoch keine Bewertungen
- Historical AntecedentsDokument3 SeitenHistorical AntecedentsLEILA PATRICIA SOSANoch keine Bewertungen
- DissertationDokument109 SeitenDissertationSayan BanerjeeNoch keine Bewertungen
- Current Research Areas - Psychology FacultyDokument6 SeitenCurrent Research Areas - Psychology FacultyAliya MojicaNoch keine Bewertungen
- Diagnostics 11 00459 v2Dokument15 SeitenDiagnostics 11 00459 v2Gabriella EvitaNoch keine Bewertungen
- Bhavna KatejaDokument2 SeitenBhavna KatejaShahabaz UbaiseNoch keine Bewertungen
- Minimally Invasive Endodontics Challenging Prevailing Paradigms PDFDokument7 SeitenMinimally Invasive Endodontics Challenging Prevailing Paradigms PDFLina SaadNoch keine Bewertungen
- Chronic Interstitial Nephritis in Agricultural.20Dokument8 SeitenChronic Interstitial Nephritis in Agricultural.20corazones.232.mgNoch keine Bewertungen
- Alana Fournet - Radiant Powerful You-Intentional Health For WomenDokument20 SeitenAlana Fournet - Radiant Powerful You-Intentional Health For Womenlawrece_acc5589Noch keine Bewertungen
- Tahun Nomor NamaDokument18 SeitenTahun Nomor NamaALDO STEFANUZ (STIK)Noch keine Bewertungen
- Hypertension Protocol Mukta VatiDokument18 SeitenHypertension Protocol Mukta VatiDr. Avnish UpadhyayNoch keine Bewertungen
- Biomecanica Da ObesidadeDokument9 SeitenBiomecanica Da ObesidadehgtrainerNoch keine Bewertungen
- Study Notes 3Dokument2 SeitenStudy Notes 3AfriantoNoch keine Bewertungen
- Research On Drug Abuse in St. HubertDokument16 SeitenResearch On Drug Abuse in St. HubertSt. Gabby100% (1)
- Mapeh ReportDokument10 SeitenMapeh ReportMikang LorNoch keine Bewertungen
- Group 1 - Posterior Disk Bulge at L5-S1 With Bilateral Severe Neural Canal Narrowing PDFDokument7 SeitenGroup 1 - Posterior Disk Bulge at L5-S1 With Bilateral Severe Neural Canal Narrowing PDFKatNoch keine Bewertungen
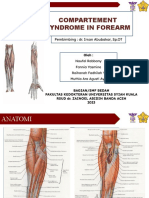
- Forearm Compartment SyndromeDokument19 SeitenForearm Compartment SyndromenaufalNoch keine Bewertungen
- Mri Lumbar Spine: Clinical Info: Technique: FindingsDokument2 SeitenMri Lumbar Spine: Clinical Info: Technique: FindingsshoaibNoch keine Bewertungen
- Lingual-Keys For SuccessDokument8 SeitenLingual-Keys For SuccessGoutam NookalaNoch keine Bewertungen
- DSM 5 Ocd ChapterDokument3 SeitenDSM 5 Ocd ChapterIva IzgubilaprezimeNoch keine Bewertungen
- ICR Wa WEBDokument8 SeitenICR Wa WEBไกรสร เต็งNoch keine Bewertungen
- Indivior PLC Presents at Morgan Stanley 19th Annual Global Healthcare Conference, Sep-13-2021 09 - 30 AMDokument12 SeitenIndivior PLC Presents at Morgan Stanley 19th Annual Global Healthcare Conference, Sep-13-2021 09 - 30 AMKing KingssonNoch keine Bewertungen
- Learning Objectives: Nstp-Cwts I Module 1 - Drug Education in The PhilippinesDokument13 SeitenLearning Objectives: Nstp-Cwts I Module 1 - Drug Education in The PhilippinesALMONIA, GEZZIEL GANIRNoch keine Bewertungen
- Acute PainDokument4 SeitenAcute PainCuttie Anne GalangNoch keine Bewertungen
- CS Drug Study NotesDokument11 SeitenCS Drug Study NotestwnynyiskaNoch keine Bewertungen
- Diagnostic Report: FinalDokument2 SeitenDiagnostic Report: FinalRAGHA DIAGNOSTIC CENTRENoch keine Bewertungen