Das könnte Ihnen auch gefallen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (894)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (587)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (119)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2219)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- SECOND PERIODICAL TEST in TLE 9Dokument3 SeitenSECOND PERIODICAL TEST in TLE 9Lima Alpha91% (103)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- Effects of Sucrose Concentration On Cell Respiration in YeastDokument7 SeitenEffects of Sucrose Concentration On Cell Respiration in YeastRachel Utomo83% (23)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- PhysicsDokument4 SeitenPhysicsAlaa Mohyeldin47% (17)
- AC7101.1 Rev G 2Dokument37 SeitenAC7101.1 Rev G 2Namelezz ShadowwNoch keine Bewertungen
- CASE Study PTBDokument53 SeitenCASE Study PTBmeleanaquino94% (16)
- Kathrein Antenna Dual BandDokument4 SeitenKathrein Antenna Dual BandAmine AchrafNoch keine Bewertungen
- ACD NMR ProcessingDokument3 SeitenACD NMR ProcessingAlaa MohyeldinNoch keine Bewertungen
- Inertia Tensor and Cross Product In: N-Dimensions SpaceDokument7 SeitenInertia Tensor and Cross Product In: N-Dimensions SpaceAlaa MohyeldinNoch keine Bewertungen
- Dynamics and Relativity Course OverviewDokument167 SeitenDynamics and Relativity Course Overviewopenid_AePkLAJcNoch keine Bewertungen
- Lecture 07Dokument22 SeitenLecture 07Ashim DuttaNoch keine Bewertungen
- Extract Pages From (George B. Arfken, Hans J.Dokument3 SeitenExtract Pages From (George B. Arfken, Hans J.Alaa MohyeldinNoch keine Bewertungen
- 12 Damped Harmonic OscillatorDokument3 Seiten12 Damped Harmonic OscillatorBora CanbulaNoch keine Bewertungen
- 008.5 Intro To Dynamical SystemsDokument23 Seiten008.5 Intro To Dynamical SystemsAlaa MohyeldinNoch keine Bewertungen
- The Multifunctional BIOCHIP: Duke UniversityDokument1 SeiteThe Multifunctional BIOCHIP: Duke UniversityAlaa MohyeldinNoch keine Bewertungen
- Crash Course On VectorsDokument40 SeitenCrash Course On VectorsjdoflaNoch keine Bewertungen
- 12 Damped Harmonic OscillatorDokument3 Seiten12 Damped Harmonic OscillatorBora CanbulaNoch keine Bewertungen
- Human Exposure To Mobile Phone EM RadiationDokument4 SeitenHuman Exposure To Mobile Phone EM RadiationAlaa MohyeldinNoch keine Bewertungen
- s2017 ActivitiesDokument0 Seitens2017 ActivitiesAlaa MohyeldinNoch keine Bewertungen
- Ielts Speaking Part One Dice Game and Question BrainstormingDokument3 SeitenIelts Speaking Part One Dice Game and Question BrainstormingAlaa MohyeldinNoch keine Bewertungen
- Study Notes Episode 22: Taliking About Holidays and Leisure TimeDokument0 SeitenStudy Notes Episode 22: Taliking About Holidays and Leisure TimeAlaa MohyeldinNoch keine Bewertungen
- Rotation MI and TorqueDokument22 SeitenRotation MI and TorqueAlaa MohyeldinNoch keine Bewertungen
- The Power of Negative Thinking - Science-2013-Couzin-Frankel-68-9 PDFDokument2 SeitenThe Power of Negative Thinking - Science-2013-Couzin-Frankel-68-9 PDFAlaa MohyeldinNoch keine Bewertungen
- Reading Skills and Question Types: Ielts TipDokument0 SeitenReading Skills and Question Types: Ielts TipAlaa MohyeldinNoch keine Bewertungen
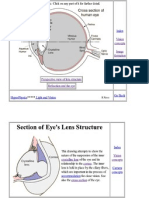
- The Human EyeDokument2 SeitenThe Human EyeAlaa MohyeldinNoch keine Bewertungen
- Trigonometric Arld Hyperbolic Iderltities: I I I IDokument4 SeitenTrigonometric Arld Hyperbolic Iderltities: I I I IAlaa MohyeldinNoch keine Bewertungen
- OCTDokument6 SeitenOCTajitNoch keine Bewertungen
- Basic Principles of Lasers: Structural Components, Properties, SafetyDokument4 SeitenBasic Principles of Lasers: Structural Components, Properties, SafetyAlaa Mohyeldin50% (2)
- InTech-Photonic Crystal Fiber For Medical ApplicationsDokument19 SeitenInTech-Photonic Crystal Fiber For Medical ApplicationsAlaa MohyeldinNoch keine Bewertungen
- Supplementary Feeding ProgramDokument2 SeitenSupplementary Feeding ProgramVictor Anthony CuaresmaNoch keine Bewertungen
- Everything You Need to Know About VodkaDokument4 SeitenEverything You Need to Know About Vodkaudbhav786Noch keine Bewertungen
- Penyakit Palpebra Dan AdneksaDokument39 SeitenPenyakit Palpebra Dan AdneksaayucicuNoch keine Bewertungen
- FICHA TECNICA Smart-EvoDokument4 SeitenFICHA TECNICA Smart-EvoClaudio A.Noch keine Bewertungen
- M96SC05 Oleo StrutDokument6 SeitenM96SC05 Oleo Strutchaumont12345Noch keine Bewertungen
- Assignment On Types of Retail Marketing: Submitted By: MR - Swapnil S. Ghag. Roll No.10 (A)Dokument9 SeitenAssignment On Types of Retail Marketing: Submitted By: MR - Swapnil S. Ghag. Roll No.10 (A)Swapnil Ghag100% (1)
- Form16 (2021-2022)Dokument2 SeitenForm16 (2021-2022)Anushka PoddarNoch keine Bewertungen
- (Distracted Subjects) CHAPTER 2. Reading The Language of Distraction - Hamlet, Macbeth, King LearDokument23 Seiten(Distracted Subjects) CHAPTER 2. Reading The Language of Distraction - Hamlet, Macbeth, King LearLCAP ConsultingNoch keine Bewertungen
- Evaluation of Personal Dust Exposure of The Rice Mill Workers in AssamDokument10 SeitenEvaluation of Personal Dust Exposure of The Rice Mill Workers in AssamIJAMTESNoch keine Bewertungen
- Physical Science Summative Exam. First QuarterDokument5 SeitenPhysical Science Summative Exam. First QuarterIsagani WagisNoch keine Bewertungen
- Technical Bulletins Miglow Underwater12Dokument1 SeiteTechnical Bulletins Miglow Underwater12Arnaud PoliNoch keine Bewertungen
- Reflexes Guide: 20+ Human Reflexes ExplainedDokument37 SeitenReflexes Guide: 20+ Human Reflexes ExplainedSalman KhanNoch keine Bewertungen
- Amnesia With Focus On Post Traumatic AmnesiaDokument27 SeitenAmnesia With Focus On Post Traumatic AmnesiaWilliam ClemmonsNoch keine Bewertungen
- B-64483EN-2 - 05 - 01 Dual Check Safety - CONNECTION MANUAL - Password - RemovedDokument270 SeitenB-64483EN-2 - 05 - 01 Dual Check Safety - CONNECTION MANUAL - Password - Removedretox9000Noch keine Bewertungen
- DaloDokument2 SeitenDalojosua tuisawauNoch keine Bewertungen
- Asian Organized CrimeDokument17 SeitenAsian Organized CrimeMagr EscaNoch keine Bewertungen
- TLC Analysis of Siling Labuyo Extract ComponentsDokument3 SeitenTLC Analysis of Siling Labuyo Extract ComponentsJan Eric Balete0% (1)
- Un Primer Acercamiento A La Escritura Científica: OPI LabDokument66 SeitenUn Primer Acercamiento A La Escritura Científica: OPI LabLolaNoch keine Bewertungen
- Extraction and Isolation of Saponins PDFDokument2 SeitenExtraction and Isolation of Saponins PDFMikeNoch keine Bewertungen
- Summative Test in Respiratory SystemDokument2 SeitenSummative Test in Respiratory SystemEden Manlosa100% (4)
- Gimnazjum Exam Practice GuideDokument74 SeitenGimnazjum Exam Practice GuideVaserd MoasleNoch keine Bewertungen
- Section III - Topic 3Dokument7 SeitenSection III - Topic 3KARINE HOVSEPYANNoch keine Bewertungen
- Common Safety Method GuidanceDokument66 SeitenCommon Safety Method GuidanceDiego UngerNoch keine Bewertungen
- Fuel Cell HandbookDokument352 SeitenFuel Cell HandbookHamza SuljicNoch keine Bewertungen
- DepEd Region I Summative Test in Cookery 9Dokument2 SeitenDepEd Region I Summative Test in Cookery 9Jessel Mejia OnzaNoch keine Bewertungen