Das könnte Ihnen auch gefallen
- Pregunta 4 (Taller No Alimentaria)Dokument6 SeitenPregunta 4 (Taller No Alimentaria)Liseth SierraNoch keine Bewertungen
- Color AntesDokument8 SeitenColor AntesBrayan SosaNoch keine Bewertungen
- Pigmentos Vegetales.Dokument4 SeitenPigmentos Vegetales.Heber RojasNoch keine Bewertungen
- Tarea 2Dokument15 SeitenTarea 2tescamillajaimesNoch keine Bewertungen
- Separación de CarotenosDokument6 SeitenSeparación de Carotenosyady1702Noch keine Bewertungen
- Quinonas y Derivados AntracenicosDokument26 SeitenQuinonas y Derivados AntracenicosYonathan Arestigue100% (2)
- Trabajo de CarotenoidesDokument5 SeitenTrabajo de CarotenoidesRonald BustamanteNoch keine Bewertungen
- AntocinaninasDokument6 SeitenAntocinaninasYesenia BPNoch keine Bewertungen
- Pigmentos NaturalesDokument18 SeitenPigmentos NaturalesAndres RequenaNoch keine Bewertungen
- ESTABILIDAD DE PIGMENTOS Exposicion QuimicaDokument29 SeitenESTABILIDAD DE PIGMENTOS Exposicion QuimicaJunior VelasquezNoch keine Bewertungen
- Conferencia 3 - CarotenoidesDokument11 SeitenConferencia 3 - CarotenoidesDianelis Alonso LeyvaNoch keine Bewertungen
- Los PigmentosDokument20 SeitenLos PigmentosRosi Briceño QuinteroNoch keine Bewertungen
- Pigmentos AccesoriosDokument3 SeitenPigmentos AccesoriosGuadalupe Lara50% (4)
- CarotenoideDokument6 SeitenCarotenoideshady182100% (1)
- CarotenoidesDokument10 SeitenCarotenoidesJose Armando Herrera MendozaNoch keine Bewertungen
- Experimento de Extraccion de Pigmentos de Las PlantasDokument12 SeitenExperimento de Extraccion de Pigmentos de Las PlantasJose Manuel Benites PalomaresNoch keine Bewertungen
- Practica Por Coloracion.Dokument12 SeitenPractica Por Coloracion.Dalessia Tello CamposNoch keine Bewertungen
- Seminario Fisio Vegetal AntocianinasDokument11 SeitenSeminario Fisio Vegetal AntocianinasCarlos Carretero100% (1)
- RodofitosDokument3 SeitenRodofitosgrupoquimicaNoch keine Bewertungen
- Clorofila y Sus PigmentosDokument4 SeitenClorofila y Sus PigmentosCarlos A. ValdezNoch keine Bewertungen
- Flavonoides de RosasDokument10 SeitenFlavonoides de RosasOrana Sarmiento Sánchez100% (1)
- Carotenoides FinalDokument40 SeitenCarotenoides FinalRoux MarrerosNoch keine Bewertungen
- Sesion 14 Quimica Organica IiiDokument35 SeitenSesion 14 Quimica Organica IiiGiampierre VillenaNoch keine Bewertungen
- CARBOHIDRATOSDokument30 SeitenCARBOHIDRATOSJuventusNoch keine Bewertungen
- Los FlavonoidesDokument2 SeitenLos FlavonoidesMaria Del Mar Hernández GiraldoNoch keine Bewertungen
- AlcaloidesDokument31 SeitenAlcaloideshellpawn00Noch keine Bewertungen
- GLICÓSIDODokument19 SeitenGLICÓSIDOElmer Oliva71% (7)
- Sesion 14 TerpenoidesDokument36 SeitenSesion 14 TerpenoidesBeatriz MarquezNoch keine Bewertungen
- Extraccion de CarotenosDokument13 SeitenExtraccion de CarotenosceciliaNoch keine Bewertungen
- Clasificación de Colorantes Qm-Tc-Jenny Trinidad Paricahua Pari IV SemestreDokument4 SeitenClasificación de Colorantes Qm-Tc-Jenny Trinidad Paricahua Pari IV SemestreSumita Sanchez MamaniNoch keine Bewertungen
- AlcaloidesDokument8 SeitenAlcaloidesChristian J. Rosas JarquínNoch keine Bewertungen
- PN Informe 9Dokument3 SeitenPN Informe 9Jaqueline CruzNoch keine Bewertungen
- Temperstura 60 Extraccion de FlavonoidesDokument14 SeitenTemperstura 60 Extraccion de Flavonoidesjuan carlosNoch keine Bewertungen
- Compuestos Fenolicos 3Dokument86 SeitenCompuestos Fenolicos 3Jaime Alejandro Godinez FloresNoch keine Bewertungen
- Clase 2 - Carotenos y FotosintesisDokument17 SeitenClase 2 - Carotenos y Fotosintesistaco inversionesNoch keine Bewertungen
- Color, Aroma y SaborDokument79 SeitenColor, Aroma y Saborelver regalado goicocheaNoch keine Bewertungen
- Identificación de Alcaloides FarmacognDokument6 SeitenIdentificación de Alcaloides FarmacognJESSICA LILI MATIAS VARGASNoch keine Bewertungen
- AlcaloidesDokument31 SeitenAlcaloidesStln AlexisNoch keine Bewertungen
- CarotenoideDokument9 SeitenCarotenoideSAntonio RamírezNoch keine Bewertungen
- CAROTENOIDESDokument15 SeitenCAROTENOIDESPrinjackNoch keine Bewertungen
- VacuolasDokument12 SeitenVacuolasMarco Rafael DíazNoch keine Bewertungen
- Practica 2 13oDokument6 SeitenPractica 2 13oAlfredo TorresgNoch keine Bewertungen
- Carbohidratos - 55Dokument87 SeitenCarbohidratos - 55Michael Jhonatan100% (1)
- Trabajo Especial FitogeneticaDokument27 SeitenTrabajo Especial Fitogeneticafelixmas007Noch keine Bewertungen
- PigmentosDokument7 SeitenPigmentosSofia Salirrosas ZapataNoch keine Bewertungen
- Terpenos Presentacion Final2Dokument43 SeitenTerpenos Presentacion Final2Jekk MatiNoch keine Bewertungen
- VacuolasDokument12 SeitenVacuolasCharito Rojas AlbarránNoch keine Bewertungen
- Clasificación Plantas Medicinales Por Nombre Común LauraDokument10 SeitenClasificación Plantas Medicinales Por Nombre Común LauraMary AnguloNoch keine Bewertungen
- Antocianina - PDF 9Dokument9 SeitenAntocianina - PDF 9luiggi77Noch keine Bewertungen
- Carbohidratos 1Dokument31 SeitenCarbohidratos 1Sisley FilosNoch keine Bewertungen
- Antraquinonicos Drogas Con GlicosidosDokument4 SeitenAntraquinonicos Drogas Con GlicosidosCARLOS USCANoch keine Bewertungen
- 1 C SaponinasDokument44 Seiten1 C SaponinasBetzaida Vilca Mamani100% (2)
- Informe 3Dokument7 SeitenInforme 3Odalis De SheeranNoch keine Bewertungen
- Marco Teórico FotosintesisDokument3 SeitenMarco Teórico Fotosintesislalala lal0% (1)
- Quimiotaxonomía FitoquimicaDokument21 SeitenQuimiotaxonomía FitoquimicaJhon R. Aquino60% (5)
- Extraccion de Colorantes 402 - para Expo DefinitivaDokument40 SeitenExtraccion de Colorantes 402 - para Expo DefinitivaJanina Muñoz de SantanaNoch keine Bewertungen
- Metabolitos secundarios presentes en algunas plantas de ColombiaVon EverandMetabolitos secundarios presentes en algunas plantas de ColombiaNoch keine Bewertungen
- Las 100 Empresas Mas Grandes Del Valle Del CaucaDokument10 SeitenLas 100 Empresas Mas Grandes Del Valle Del CaucaMauricioNoch keine Bewertungen
- Leyes Energeticas de La FragmentacionDokument5 SeitenLeyes Energeticas de La FragmentacionMauricioNoch keine Bewertungen
- Equilibrio Metanol AguaDokument1 SeiteEquilibrio Metanol AguaMauricioNoch keine Bewertungen
- Higiene y Seguridad IndustrialDokument2 SeitenHigiene y Seguridad IndustrialMauricio0% (1)
- Tabla de Propiedades Del Agua (Unidades SI)Dokument10 SeitenTabla de Propiedades Del Agua (Unidades SI)MauricioNoch keine Bewertungen
- IrosoDokument13 SeitenIrosoFabianoNoch keine Bewertungen
- Renovación, Migración y Diferenciación de Las CélulasDokument15 SeitenRenovación, Migración y Diferenciación de Las CélulasAlejandro Chung BenitesNoch keine Bewertungen
- 16 Si Diagnostico PrenatalDokument7 Seiten16 Si Diagnostico PrenatalSephiroth_Kun_4Noch keine Bewertungen
- Alimentos TransgenicosDokument22 SeitenAlimentos TransgenicosKevinteru -kunNoch keine Bewertungen
- Escuelas de CantoDokument3 SeitenEscuelas de CantoGabriela Belén Ojeda SilvaNoch keine Bewertungen
- Pie PronadorDokument14 SeitenPie PronadorOmar RamírezNoch keine Bewertungen
- El Ciclo CelularDokument4 SeitenEl Ciclo CelularDaiana AlmadaNoch keine Bewertungen
- Clave Dicotómica VertebradosDokument4 SeitenClave Dicotómica VertebradosAntonio Lloret50% (2)
- Flavonoides y Aloe Vera InfDokument7 SeitenFlavonoides y Aloe Vera InfAlejandra HoyosNoch keine Bewertungen
- Plomo, Mercurio y ArsenicoDokument8 SeitenPlomo, Mercurio y ArsenicoJeimmy MartínezNoch keine Bewertungen
- Sesion Ciencia 04 de JulioDokument5 SeitenSesion Ciencia 04 de JulioZonya Fer100% (1)
- Normas Básicas de Higiene y Seguridad IndustrialDokument5 SeitenNormas Básicas de Higiene y Seguridad IndustrialDavid Fernando Vargas Blanco100% (1)
- Trastornos de La Migración NeuronalDokument3 SeitenTrastornos de La Migración NeuronalAlfredo TovarNoch keine Bewertungen
- Sindrome de Ovario PoliquisticoDokument3 SeitenSindrome de Ovario PoliquisticoRosana LujanNoch keine Bewertungen
- Estudio Violencia en Tres Poblaciones de Mujeres en BoliviaDokument225 SeitenEstudio Violencia en Tres Poblaciones de Mujeres en BoliviaGracia Violeta Ross QuirogaNoch keine Bewertungen
- Caros en Cuyes y AvesDokument8 SeitenCaros en Cuyes y AvesGIAN VICTOR ARO QUISPENoch keine Bewertungen
- Carta de Brigadistas MASSALADokument5 SeitenCarta de Brigadistas MASSALASEGURIDAD Y SALUD EN EL TRABAJONoch keine Bewertungen
- 5 Oncología - Schwartz. Principios de Cirugía, 11e - AccessMedicina - McGraw Hill MedicalDokument15 Seiten5 Oncología - Schwartz. Principios de Cirugía, 11e - AccessMedicina - McGraw Hill MedicalAguero AgnisNoch keine Bewertungen
- Tutoria 5 y 6 UeesDokument7 SeitenTutoria 5 y 6 UeesYadi FiencoNoch keine Bewertungen
- Manzanares - ConcepcionDokument26 SeitenManzanares - ConcepcionBrayanKennethNoch keine Bewertungen
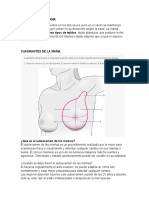
- Anatomia de La MamaDokument8 SeitenAnatomia de La MamaninyNoch keine Bewertungen
- ENSAYO El Sistema Nacional de Salud en MexDokument9 SeitenENSAYO El Sistema Nacional de Salud en MexEduardo Nava0% (1)
- Pruebas Selectivas 2023 Cuaderno de ExamenDokument74 SeitenPruebas Selectivas 2023 Cuaderno de ExamennitrolocusNoch keine Bewertungen
- Funciones Del Sistema UrinarioDokument3 SeitenFunciones Del Sistema UrinarioChabelaHuertasCrispin100% (1)
- Via ParenteralDokument4 SeitenVia Parenteralseguro socialNoch keine Bewertungen
- Poda de UvaDokument1 SeitePoda de UvaKarinaRscNoch keine Bewertungen
- Programa Internado Obstetricia y Ginecologia 2012Dokument24 SeitenPrograma Internado Obstetricia y Ginecologia 2012Cuno CarinaNoch keine Bewertungen
- FGHDGHDokument7 SeitenFGHDGHGabriela ChavezNoch keine Bewertungen
- Control Prenatal y Anemia en El EmbarazoDokument10 SeitenControl Prenatal y Anemia en El EmbarazoTatiana OchoaNoch keine Bewertungen
- Guía Didáctica Manejo Integrado de PlagasDokument41 SeitenGuía Didáctica Manejo Integrado de Plagaspercy vega villasanteNoch keine Bewertungen