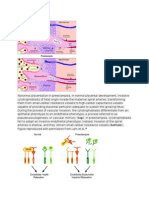
Das könnte Ihnen auch gefallen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (894)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (587)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (119)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2219)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Nursing Fundamentals Exam ReviewDokument10 SeitenNursing Fundamentals Exam ReviewAlain CheryNoch keine Bewertungen
- Toxicology ReportDokument2 SeitenToxicology Reportapi-457390058Noch keine Bewertungen
- Definition of HypoglycemiaDokument9 SeitenDefinition of HypoglycemiaAchiApsariNoch keine Bewertungen
- Jurnal KulitDokument12 SeitenJurnal KulitAulia FitrianiNoch keine Bewertungen
- Abnormal Placentation in PreeclampsiaDokument6 SeitenAbnormal Placentation in PreeclampsiaAchiApsariNoch keine Bewertungen
- Wart 2011 Efficacy of Intralesional BleomycinDokument4 SeitenWart 2011 Efficacy of Intralesional BleomycinAchiApsariNoch keine Bewertungen
- Abnormal Placentation in PreeclampsiaDokument6 SeitenAbnormal Placentation in PreeclampsiaAchiApsariNoch keine Bewertungen
- Acute Morbidity Associated With Scabies and OthDokument7 SeitenAcute Morbidity Associated With Scabies and OthAchiApsariNoch keine Bewertungen
- Gummin2018 PDFDokument204 SeitenGummin2018 PDFAelton AraujoNoch keine Bewertungen
- EphedraDokument9 SeitenEphedraapi-367367047Noch keine Bewertungen
- RSVDokument8 SeitenRSVilyas9558Noch keine Bewertungen
- Paediatric History Collection FormatDokument7 SeitenPaediatric History Collection FormatSathya Palanisamy100% (1)
- Conventional RadiologyDokument2 SeitenConventional RadiologyPier Cedron LescanoNoch keine Bewertungen
- Disease Doesn't Occur in A Vacuum Disease Is Not Randomly Distributed - Epidemiology Uses Systematic Approach ToDokument44 SeitenDisease Doesn't Occur in A Vacuum Disease Is Not Randomly Distributed - Epidemiology Uses Systematic Approach Toاسامة محمد السيد رمضانNoch keine Bewertungen
- Genitourinary ReviewDokument103 SeitenGenitourinary ReviewGideon P. CasasNoch keine Bewertungen
- Chronic and Acute GoutDokument20 SeitenChronic and Acute GoutDan TranNoch keine Bewertungen
- Chronic Kidney Disease in Homoeopathy Part-1Dokument31 SeitenChronic Kidney Disease in Homoeopathy Part-1Tarique ImamNoch keine Bewertungen
- Chapter 03: Health Policy and The Delivery SystemDokument11 SeitenChapter 03: Health Policy and The Delivery SystemStephany ZatarainNoch keine Bewertungen
- Diabetes in Pregnancy: Management ofDokument8 SeitenDiabetes in Pregnancy: Management ofdiana rossNoch keine Bewertungen
- Parenteral Nutrition CalculationsDokument62 SeitenParenteral Nutrition CalculationsaqeelaadnanNoch keine Bewertungen
- Aa 11Dokument5 SeitenAa 11Eriekafebriayana RNoch keine Bewertungen
- Antibiotic Skin TestingDokument8 SeitenAntibiotic Skin TestingFitz JaminitNoch keine Bewertungen
- Hipocrates - VOLUME 1Dokument452 SeitenHipocrates - VOLUME 1Heitor Murillo CarnioNoch keine Bewertungen
- Chronic Gastritis and Peptic Ulcer Disease GuideDokument32 SeitenChronic Gastritis and Peptic Ulcer Disease GuideDian Ekawati100% (2)
- Drug Study (Newborn Screening)Dokument2 SeitenDrug Study (Newborn Screening)Aaron Christian AlmoiteNoch keine Bewertungen
- Biological Child PsychiatryDokument264 SeitenBiological Child Psychiatrydejoguna126100% (2)
- Sutter Health may cancel up to 90k vaccine appointments due to low supplyDokument2 SeitenSutter Health may cancel up to 90k vaccine appointments due to low supplyAryan PgzNoch keine Bewertungen
- Manual Reprocessing Final Version 2.0Dokument29 SeitenManual Reprocessing Final Version 2.0Enaj AzilgNoch keine Bewertungen
- NHSRC - Operational Guidelines On Maternal and Newborn HealthDokument46 SeitenNHSRC - Operational Guidelines On Maternal and Newborn HealthNational Child Health Resource Centre (NCHRC)Noch keine Bewertungen
- Protocols Sepsis Treatment Stony BrookDokument6 SeitenProtocols Sepsis Treatment Stony BrookVicky Chrystine SianiparNoch keine Bewertungen
- Discharge PlanDokument4 SeitenDischarge PlanPaul Loujin LeeNoch keine Bewertungen
- Kelompok 7 - 4b - Buku Saku Antibiotik Dan AntivirusDokument62 SeitenKelompok 7 - 4b - Buku Saku Antibiotik Dan AntivirusWidya PratiwiNoch keine Bewertungen
- Dengue Hemorrhagic ShitDokument28 SeitenDengue Hemorrhagic ShitRebecca FabonNoch keine Bewertungen
- Peripheral IntravenousDokument12 SeitenPeripheral IntravenousDewi List0% (1)
- Tinnitus Today December 1998 Vol 23, No 4Dokument29 SeitenTinnitus Today December 1998 Vol 23, No 4American Tinnitus AssociationNoch keine Bewertungen
- WVSU Adult Nursing ProcessDokument9 SeitenWVSU Adult Nursing ProcessMark ArconadaNoch keine Bewertungen