Das könnte Ihnen auch gefallen
- Monoraphid and Naviculoid Diatoms from the Coastal Laurentian Great LakesVon EverandMonoraphid and Naviculoid Diatoms from the Coastal Laurentian Great LakesAndrzej WitkowskiNoch keine Bewertungen
- Jurnal CephalochordataDokument3 SeitenJurnal CephalochordataAnna KareninaNoch keine Bewertungen
- FSR 175Dokument10 SeitenFSR 175Michaiel PiticarNoch keine Bewertungen
- Stamou2017 Article TaxonomicDistinctnessIndicesFoDokument13 SeitenStamou2017 Article TaxonomicDistinctnessIndicesFoCristian VargasNoch keine Bewertungen
- Rhodes 2010Dokument11 SeitenRhodes 2010AnghelloLozanoDíazNoch keine Bewertungen
- Independant Study ReportDokument12 SeitenIndependant Study Reportapi-659954273Noch keine Bewertungen
- Stable Isotopes Indicate Individual Level Trophic Diversity in Freshwater GastropodDokument5 SeitenStable Isotopes Indicate Individual Level Trophic Diversity in Freshwater GastropodMarco Antonio SolisNoch keine Bewertungen
- Diversity and Antimicrobial Activities of Surface-Attached Marine Bacteria From Sydney Harbour, AustraliaDokument12 SeitenDiversity and Antimicrobial Activities of Surface-Attached Marine Bacteria From Sydney Harbour, Australiadaie90Noch keine Bewertungen
- tmp2F18 TMPDokument13 Seitentmp2F18 TMPFrontiersNoch keine Bewertungen
- Camargo Et Al 2004Dokument10 SeitenCamargo Et Al 2004rini susilowatiNoch keine Bewertungen
- Shallow Lakes 2002Dokument309 SeitenShallow Lakes 2002testvietnamNoch keine Bewertungen
- Biola 2017 PDFDokument12 SeitenBiola 2017 PDFBob AnggaraNoch keine Bewertungen
- Trophic State of An Urban Coastal LagoonDokument12 SeitenTrophic State of An Urban Coastal LagoonJurandir MarkanteNoch keine Bewertungen
- Proceedings of The Fifth International Conference of Research and Development, Hotel Residence The Carlos, 20-25th November, 2011, Buea, Cameroun, Published by The Pan-African Book Company 3 (2) :5-11Dokument9 SeitenProceedings of The Fifth International Conference of Research and Development, Hotel Residence The Carlos, 20-25th November, 2011, Buea, Cameroun, Published by The Pan-African Book Company 3 (2) :5-11krithideepNoch keine Bewertungen
- AlkenonasDokument6 SeitenAlkenonaskmiloNoch keine Bewertungen
- Jurnalnya Kawanku TersayangDokument13 SeitenJurnalnya Kawanku TersayangMoch Lazuardy IslamiNoch keine Bewertungen
- Pasquaud Et Al 20081Dokument13 SeitenPasquaud Et Al 20081MyrubicRubicNoch keine Bewertungen
- Ambroso Et Al 2016. Ophiuroid Biodiversity Patterns Along The Antarctic PeninsulaDokument16 SeitenAmbroso Et Al 2016. Ophiuroid Biodiversity Patterns Along The Antarctic Peninsulabyhz7wwdm4Noch keine Bewertungen
- Microbial Diversity of Boron-Rich Volcanic Hot Springs of St. Lucia, Lesser AntillesDokument11 SeitenMicrobial Diversity of Boron-Rich Volcanic Hot Springs of St. Lucia, Lesser AntillesSandip AjmireNoch keine Bewertungen
- Vertical Shifts of Particle-Attached and Free-Living Prokaryotes in Water Column Above Cold Seeps South China SeaDokument11 SeitenVertical Shifts of Particle-Attached and Free-Living Prokaryotes in Water Column Above Cold Seeps South China SeaAndrea EscobarNoch keine Bewertungen
- Aspects of The Geomorphology and Limnology of Some Molluscinhabited Freshwater Bodies in Northern NigeriaDokument10 SeitenAspects of The Geomorphology and Limnology of Some Molluscinhabited Freshwater Bodies in Northern NigeriaIOSRjournalNoch keine Bewertungen
- Grupos Funcionales Del Planton Frente Al Puerto Pesquero de AnconcitoDokument17 SeitenGrupos Funcionales Del Planton Frente Al Puerto Pesquero de AnconcitoCear Andrade RuízNoch keine Bewertungen
- Stable Isotopes of Nitrogen and Carbon in An Aquatic Food Web Recently Invaded by Dreissena (Pallas)Dokument6 SeitenStable Isotopes of Nitrogen and Carbon in An Aquatic Food Web Recently Invaded by Dreissena (Pallas)mikamirNoch keine Bewertungen
- Benitez-Villalobos Et Al. 2012Dokument10 SeitenBenitez-Villalobos Et Al. 2012Juan Manuel Jesús Guerrero QuitoránNoch keine Bewertungen
- 1 s2.0 S0048969723018752 MainDokument13 Seiten1 s2.0 S0048969723018752 MainPedroHenriqueNoch keine Bewertungen
- Aquatic Biology Annotated Bibliographies 12.16.17Dokument5 SeitenAquatic Biology Annotated Bibliographies 12.16.17Cher NicoNoch keine Bewertungen
- Structure and Diversity of Zooplankton Community in Taabo Reservoir (Cote Divoire)Dokument17 SeitenStructure and Diversity of Zooplankton Community in Taabo Reservoir (Cote Divoire)IJAR JOURNALNoch keine Bewertungen
- 17 Full PDFDokument9 Seiten17 Full PDFPriscilia YuniarNoch keine Bewertungen
- MésoplanctonsDokument34 SeitenMésoplanctonserraiouiNoch keine Bewertungen
- Pollution Shapes The Microbial Communities in River Water and SedimentsDokument23 SeitenPollution Shapes The Microbial Communities in River Water and Sedimentsleloandiswa2Noch keine Bewertungen
- Diversity of PlanktonDokument10 SeitenDiversity of Planktonsara unbeknaNoch keine Bewertungen
- Borell Et AlDokument8 SeitenBorell Et Alapi-3828346Noch keine Bewertungen
- Lipsewersetal 2020Dokument18 SeitenLipsewersetal 2020Ishu NuwaNoch keine Bewertungen
- Laila Khalid Ettahiri 2014Dokument12 SeitenLaila Khalid Ettahiri 2014Boubker PouperNoch keine Bewertungen
- 6Dokument9 Seiten6JaninaZamoraNoch keine Bewertungen
- Symbiodinium Diversity Associated With ZDokument9 SeitenSymbiodinium Diversity Associated With ZThu Anh NguyenNoch keine Bewertungen
- Basin Specific Changes in Filamentous Cyanobacteria Community C - 2020 - HarmfulDokument12 SeitenBasin Specific Changes in Filamentous Cyanobacteria Community C - 2020 - Harmfulzan zibarNoch keine Bewertungen
- EPT Sampling MethodDokument13 SeitenEPT Sampling MethodForFet LifeNoch keine Bewertungen
- 8721 30727 1 PBDokument8 Seiten8721 30727 1 PBdeshie_ddsNoch keine Bewertungen
- Palaeogeography, Palaeoclimatology, PalaeoecologyDokument15 SeitenPalaeogeography, Palaeoclimatology, PalaeoecologyValeska MolinaNoch keine Bewertungen
- GJESM Volume 7 Issue 1 Pages 79-88Dokument10 SeitenGJESM Volume 7 Issue 1 Pages 79-88GJESMNoch keine Bewertungen
- TMP 2 CCDokument13 SeitenTMP 2 CCFrontiersNoch keine Bewertungen
- DiatomDokument16 SeitenDiatomRecto SutismaNoch keine Bewertungen
- Distribution and Identification of Luminous Bacteria From The Sargasso SeaDokument6 SeitenDistribution and Identification of Luminous Bacteria From The Sargasso SeajavierNoch keine Bewertungen
- Fmars 07 00575Dokument19 SeitenFmars 07 00575cpavloudNoch keine Bewertungen
- Issue02b 09 2021Dokument16 SeitenIssue02b 09 2021Vane NavarreteNoch keine Bewertungen
- Remote Estimation of Chlorophyll Concentration in Productive Waters Principals, Algorithm Development and ValidationDokument12 SeitenRemote Estimation of Chlorophyll Concentration in Productive Waters Principals, Algorithm Development and ValidationFLávio ViníciusNoch keine Bewertungen
- Elemental Proxies For Paleosalinity Analysis of Ancient Shales.. - Wei and Algeo - 2020Dokument26 SeitenElemental Proxies For Paleosalinity Analysis of Ancient Shales.. - Wei and Algeo - 2020John ManriqueNoch keine Bewertungen
- tmpB2A9 TMPDokument11 SeitentmpB2A9 TMPFrontiersNoch keine Bewertungen
- FallDokument8 SeitenFallArief DeswantaraNoch keine Bewertungen
- Spatial and Nycthemeral Variations of Zooplankton in Urban Aquatic Ecosystems of Daloa (West-Central of Cote Divoire)Dokument10 SeitenSpatial and Nycthemeral Variations of Zooplankton in Urban Aquatic Ecosystems of Daloa (West-Central of Cote Divoire)IJAR JOURNALNoch keine Bewertungen
- rDNA 16 SDokument13 SeitenrDNA 16 Sbioefl33Noch keine Bewertungen
- Aquino Et Al., 2011 - Fitoplâncton de Uma Lagoa de Estabilização No Nordeste Do Brasil PDFDokument7 SeitenAquino Et Al., 2011 - Fitoplâncton de Uma Lagoa de Estabilização No Nordeste Do Brasil PDFUbirajara FernandesNoch keine Bewertungen
- Diffuse Flow Environments Within Basalt-And Sediment-Based Hydrothermal Vent Ecosystems Harbor Specialized Microbial CommunitiesDokument15 SeitenDiffuse Flow Environments Within Basalt-And Sediment-Based Hydrothermal Vent Ecosystems Harbor Specialized Microbial CommunitiesScientificBoyNoch keine Bewertungen
- Publications Phyto List1Dokument93 SeitenPublications Phyto List1Hendry WijayantiNoch keine Bewertungen
- 2011 Alava Egg PopDokument12 Seiten2011 Alava Egg PopTammy BoavidaNoch keine Bewertungen
- Mitchelmore Et Al., (2019)Dokument13 SeitenMitchelmore Et Al., (2019)Asistente LaboratorioNoch keine Bewertungen
- Envi Toxicity in Baltic SeaDokument8 SeitenEnvi Toxicity in Baltic SeaAleine Leilanie OroNoch keine Bewertungen
- The Salinity Tolerance of Riverine Microinvertebrates From The Southern Murray-Darling BasinDokument50 SeitenThe Salinity Tolerance of Riverine Microinvertebrates From The Southern Murray-Darling BasinHassan AL-HilallyNoch keine Bewertungen
- Sampaio Et Al 2007Dokument9 SeitenSampaio Et Al 2007Bruna FacundesNoch keine Bewertungen
- Chemical Waste GNDokument14 SeitenChemical Waste GNgangsNoch keine Bewertungen
- Improved Method For CO2 MeasurementsDokument4 SeitenImproved Method For CO2 MeasurementsgangsNoch keine Bewertungen
- Guidance Chemical StorageDokument22 SeitenGuidance Chemical StoragegangsNoch keine Bewertungen
- Qualitative Analysis of Amino Acids and Proteins and Carbohydrates and Lipids and SteroidsDokument35 SeitenQualitative Analysis of Amino Acids and Proteins and Carbohydrates and Lipids and SteroidsgangsNoch keine Bewertungen
- TOCV Combustion Praxistips Eng 04IDokument35 SeitenTOCV Combustion Praxistips Eng 04IgangsNoch keine Bewertungen
- Multicomponent Spectral Fitting: ICP-Optical EmissionDokument2 SeitenMulticomponent Spectral Fitting: ICP-Optical EmissiongangsNoch keine Bewertungen
- Marine AlkenonesDokument14 SeitenMarine AlkenonesgangsNoch keine Bewertungen
- Ibms Diploma Biomedical Science Guidance NotesDokument9 SeitenIbms Diploma Biomedical Science Guidance NotesgangsNoch keine Bewertungen
- XP 1500Dokument1 SeiteXP 1500gangsNoch keine Bewertungen
- Indian Journals Indexed in Web of Science 14 2019 PDFDokument5 SeitenIndian Journals Indexed in Web of Science 14 2019 PDFRAMAKRISHNA VNoch keine Bewertungen
- Genetics: An Introduction: Peter J. RussellDokument72 SeitenGenetics: An Introduction: Peter J. RussellAli SeenaNoch keine Bewertungen
- PHD: Host-Parasite InteractionsDokument3 SeitenPHD: Host-Parasite InteractionsAlemayehu DargeNoch keine Bewertungen
- Jurnal EvolusiDokument19 SeitenJurnal EvolusiAdicahyoo SentosaNoch keine Bewertungen
- Answers To Chapter 5: (In-Text & Asterisked Problems)Dokument10 SeitenAnswers To Chapter 5: (In-Text & Asterisked Problems)平野健太朗Noch keine Bewertungen
- Importanceof CarbohydratesDokument3 SeitenImportanceof CarbohydratescraigNoch keine Bewertungen
- Cyclin A Protein Specified by Maternal MRNA in Sea Urchin Eggs That Is Destroyed at Each Cleavage DivisionDokument8 SeitenCyclin A Protein Specified by Maternal MRNA in Sea Urchin Eggs That Is Destroyed at Each Cleavage Divisionjavi0% (1)
- Fluorimeter Application Note: A10-007A: Phosphatase Assays Using Difmup As SubstrateDokument3 SeitenFluorimeter Application Note: A10-007A: Phosphatase Assays Using Difmup As SubstrateloNoch keine Bewertungen
- Oncology 101: Cancer BasicsDokument74 SeitenOncology 101: Cancer BasicsMary Rose Jose GragasinNoch keine Bewertungen
- Apoptosis: Proliferation, Differentiation and Apoptosis Are Crucial To DevelopmentDokument21 SeitenApoptosis: Proliferation, Differentiation and Apoptosis Are Crucial To DevelopmentInsyirah JohariNoch keine Bewertungen
- EMTEA Study Guide FinalDokument4 SeitenEMTEA Study Guide FinalJuan GironaNoch keine Bewertungen
- ELS Q2 Answer-Key-4 PDFDokument9 SeitenELS Q2 Answer-Key-4 PDFTest Ting100% (1)
- 17-Chapter Manuscript-477-1-10-20200501Dokument14 Seiten17-Chapter Manuscript-477-1-10-20200501nikhilkapoor1999Noch keine Bewertungen
- Immunology: Teaching ObjectivesDokument52 SeitenImmunology: Teaching ObjectivesBurhanuddin ParawalaNoch keine Bewertungen
- NSW Biology Year 11 Question and Answer Booklet 2022Dokument24 SeitenNSW Biology Year 11 Question and Answer Booklet 2022annaNoch keine Bewertungen
- AvijitDokument4 SeitenAvijitAmar RajputNoch keine Bewertungen
- Kutasi Et Al-2011-Journal of Veterinary Internal Medicine PDFDokument6 SeitenKutasi Et Al-2011-Journal of Veterinary Internal Medicine PDFAnonymous uCvzE0NVNoch keine Bewertungen
- Common Medical Abbreviations For MedTech and Gen SciDokument14 SeitenCommon Medical Abbreviations For MedTech and Gen SciRigel AntilegandoNoch keine Bewertungen
- Assurance Gds Brochure Br2694en MKDokument4 SeitenAssurance Gds Brochure Br2694en MKFatimaezzahraa BouaalamNoch keine Bewertungen
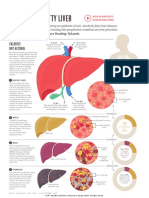
- Fatty LiverDokument2 SeitenFatty LiverKarla Edith Rodriguez Nava100% (2)
- FOOD WEB # 1: Kingdoms: Kingdom Animalia FactsDokument4 SeitenFOOD WEB # 1: Kingdoms: Kingdom Animalia Factsapi-300400997Noch keine Bewertungen
- ATEROSKLEROSISDokument9 SeitenATEROSKLEROSISNur Aini AmrunNoch keine Bewertungen
- Sathyanarayan Biochemistry 4epdfDokument35 SeitenSathyanarayan Biochemistry 4epdfAbubakar SuleimanNoch keine Bewertungen
- Amigdala HistopatologiDokument7 SeitenAmigdala HistopatologirolanNoch keine Bewertungen
- Clinical ImmunologyDokument414 SeitenClinical ImmunologyCaio LimaNoch keine Bewertungen
- MergedDokument427 SeitenMergedNishit kumar100% (1)
- Regulation of Gene ExpressionDokument20 SeitenRegulation of Gene ExpressionrainabtNoch keine Bewertungen
- Netters Essential Histology With Student Consult Access 2nd Edition Ovalle Test BankDokument35 SeitenNetters Essential Histology With Student Consult Access 2nd Edition Ovalle Test Bankjacobnigordon100% (14)
- Origami Protein HandoutDokument4 SeitenOrigami Protein HandoutRaajeswaran BaskaranNoch keine Bewertungen
- Fraga - 2005 - Epigenetic Differences Arise During The Lifetime of Mono Zygotic TwinsDokument6 SeitenFraga - 2005 - Epigenetic Differences Arise During The Lifetime of Mono Zygotic TwinstechnopsychNoch keine Bewertungen
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesVon EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesBewertung: 4.5 von 5 Sternen4.5/5 (397)
- Gut: the new and revised Sunday Times bestsellerVon EverandGut: the new and revised Sunday Times bestsellerBewertung: 4 von 5 Sternen4/5 (393)
- Why We Die: The New Science of Aging and the Quest for ImmortalityVon EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityBewertung: 4 von 5 Sternen4/5 (5)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceVon EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceBewertung: 4.5 von 5 Sternen4.5/5 (517)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisVon EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisBewertung: 3.5 von 5 Sternen3.5/5 (2)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeVon EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeNoch keine Bewertungen
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessVon Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessBewertung: 4 von 5 Sternen4/5 (33)
- Tales from Both Sides of the Brain: A Life in NeuroscienceVon EverandTales from Both Sides of the Brain: A Life in NeuroscienceBewertung: 3 von 5 Sternen3/5 (18)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsVon EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsBewertung: 4.5 von 5 Sternen4.5/5 (6)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionVon EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionBewertung: 4 von 5 Sternen4/5 (812)
- Return of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseVon EverandReturn of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseBewertung: 4.5 von 5 Sternen4.5/5 (52)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperVon EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperBewertung: 4.5 von 5 Sternen4.5/5 (16)
- Who's in Charge?: Free Will and the Science of the BrainVon EverandWho's in Charge?: Free Will and the Science of the BrainBewertung: 4 von 5 Sternen4/5 (65)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Von EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Bewertung: 4 von 5 Sternen4/5 (411)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedVon EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedBewertung: 4 von 5 Sternen4/5 (11)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomVon EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomBewertung: 4 von 5 Sternen4/5 (216)
- Good Without God: What a Billion Nonreligious People Do BelieveVon EverandGood Without God: What a Billion Nonreligious People Do BelieveBewertung: 4 von 5 Sternen4/5 (66)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorVon EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNoch keine Bewertungen
- Seven and a Half Lessons About the BrainVon EverandSeven and a Half Lessons About the BrainBewertung: 4 von 5 Sternen4/5 (110)
- Human: The Science Behind What Makes Your Brain UniqueVon EverandHuman: The Science Behind What Makes Your Brain UniqueBewertung: 3.5 von 5 Sternen3.5/5 (38)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemVon EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemBewertung: 4.5 von 5 Sternen4.5/5 (115)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldVon EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldBewertung: 4 von 5 Sternen4/5 (595)
- Minds Make Societies: How Cognition Explains the World Humans CreateVon EverandMinds Make Societies: How Cognition Explains the World Humans CreateBewertung: 4.5 von 5 Sternen4.5/5 (24)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouVon EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouBewertung: 4.5 von 5 Sternen4.5/5 (62)
- The Invention of Tomorrow: A Natural History of ForesightVon EverandThe Invention of Tomorrow: A Natural History of ForesightBewertung: 4.5 von 5 Sternen4.5/5 (5)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildVon EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildBewertung: 4.5 von 5 Sternen4.5/5 (44)