Das könnte Ihnen auch gefallen
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- What Happens in INFLAMMATION? PlayersDokument10 SeitenWhat Happens in INFLAMMATION? PlayersOdessa FileNoch keine Bewertungen
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (587)
- Efficiency of Atp ProductionDokument3 SeitenEfficiency of Atp ProductionOdessa FileNoch keine Bewertungen
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (890)
- Comments On "A Perceptual Channel For Information Transfer Over Kilometer Distances: Historical Perspective and Recent Research"Dokument3 SeitenComments On "A Perceptual Channel For Information Transfer Over Kilometer Distances: Historical Perspective and Recent Research"Odessa FileNoch keine Bewertungen
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Bionics of Spiral Structures S. v. PetukhovDokument6 SeitenBionics of Spiral Structures S. v. PetukhovOdessa FileNoch keine Bewertungen
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- 6th Central Pay Commission Salary CalculatorDokument15 Seiten6th Central Pay Commission Salary Calculatorrakhonde100% (436)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- Delayed Luminescence From Biological Systems Within The Davydov Soliton ModelDokument5 SeitenDelayed Luminescence From Biological Systems Within The Davydov Soliton ModelOdessa FileNoch keine Bewertungen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Methylation Profile PlasmaDokument4 SeitenMethylation Profile PlasmaOdessa FileNoch keine Bewertungen
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Bio Information Within The Biofield Beyond BioelectromagneticsDokument11 SeitenBio Information Within The Biofield Beyond BioelectromagneticsRobert Ri'chardNoch keine Bewertungen
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- What Is Schrodinger's NegentropyDokument15 SeitenWhat Is Schrodinger's NegentropyOdessa FileNoch keine Bewertungen
- 3 The Unexpected Capacity of Melanin To PDFDokument2 Seiten3 The Unexpected Capacity of Melanin To PDFOdessa FileNoch keine Bewertungen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Inositol Modulation of Essential Metabolic Pathways of Insulin Resistance (AugSept 2015) Townsend Letter, Alternative Medicine MagazineDokument5 SeitenInositol Modulation of Essential Metabolic Pathways of Insulin Resistance (AugSept 2015) Townsend Letter, Alternative Medicine MagazineOdessa FileNoch keine Bewertungen
- What Are Colloids... IntroductionDokument12 SeitenWhat Are Colloids... IntroductionOdessa FileNoch keine Bewertungen
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Veech03 - Therapeutic Implications of Ketone BodiesDokument11 SeitenVeech03 - Therapeutic Implications of Ketone Bodiessmtm06Noch keine Bewertungen
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Buttermilk ComponentsDokument22 SeitenButtermilk ComponentsOdessa FileNoch keine Bewertungen
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2219)
- Acid Base BalanceDokument5 SeitenAcid Base BalanceOdessa FileNoch keine Bewertungen
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Plasma - The Genesis of The Word Mario J. PinheiroDokument2 SeitenPlasma - The Genesis of The Word Mario J. PinheiroOdessa FileNoch keine Bewertungen
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Effectiveness of Chondroitin Sulphate and Other GlycosaminoglycansDokument8 SeitenEffectiveness of Chondroitin Sulphate and Other GlycosaminoglycansOdessa FileNoch keine Bewertungen
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (265)
- Water at Surfaces..Philip BallDokument3 SeitenWater at Surfaces..Philip BallOdessa FileNoch keine Bewertungen
- Evolution of Consciousness and The Emerg PDFDokument35 SeitenEvolution of Consciousness and The Emerg PDFOdessa FileNoch keine Bewertungen
- I. Bokkon and V. Salari - Information Storing by BiomagnetitesDokument17 SeitenI. Bokkon and V. Salari - Information Storing by BiomagnetitesIrokkNoch keine Bewertungen
- Autophagy - A Cell Repair Mechanism That Retards Ageing and Age-Associated Diseases and Can Be Intensified PharmacologicallyDokument8 SeitenAutophagy - A Cell Repair Mechanism That Retards Ageing and Age-Associated Diseases and Can Be Intensified PharmacologicallyOdessa FileNoch keine Bewertungen
- Therapeutic Translation of Microbial Catabolism To The LysosomeDokument8 SeitenTherapeutic Translation of Microbial Catabolism To The LysosomeOdessa FileNoch keine Bewertungen
- Alternative Splicing Rewires Cellular Metabolism To Turn On The Warburg Effect PDFDokument6 SeitenAlternative Splicing Rewires Cellular Metabolism To Turn On The Warburg Effect PDFOdessa FileNoch keine Bewertungen
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Total ORACsc and Why All 5 Radicals MatterDokument1 SeiteTotal ORACsc and Why All 5 Radicals MatterOdessa FileNoch keine Bewertungen
- 17th Annual World Congress On Anti-Aging Medicine 23-25 April 2009, Orlando, Florida, USADokument4 Seiten17th Annual World Congress On Anti-Aging Medicine 23-25 April 2009, Orlando, Florida, USAOdessa FileNoch keine Bewertungen
- Matveev-2002-Revolution and Counter Revolution PDFDokument4 SeitenMatveev-2002-Revolution and Counter Revolution PDFOdessa FileNoch keine Bewertungen
- Silybin - Vitamin E - PhospholipidsDokument2 SeitenSilybin - Vitamin E - PhospholipidsOdessa FileNoch keine Bewertungen
- Berberine - A Potential Multipotent Natural Product To Combat Alzheimer's DiseaseDokument9 SeitenBerberine - A Potential Multipotent Natural Product To Combat Alzheimer's DiseaseOdessa FileNoch keine Bewertungen
- BiflacDokument17 SeitenBiflacMohamed SafwanNoch keine Bewertungen
- NRF2 Activation LeafletDokument4 SeitenNRF2 Activation LeafletOdessa FileNoch keine Bewertungen
- Biology - Stem CellsDokument9 SeitenBiology - Stem CellsLeen ANoch keine Bewertungen
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (119)
- 0A - Revision Table Eng TRDokument2 Seiten0A - Revision Table Eng TRSpontan GezginNoch keine Bewertungen
- Lect Skin4aDokument98 SeitenLect Skin4akazchandranNoch keine Bewertungen
- Tinywow PhysicalEducation12 2022 (1) 41424952 7Dokument38 SeitenTinywow PhysicalEducation12 2022 (1) 41424952 7HwhaibNoch keine Bewertungen
- Physiology of The Gastrointestinal Tract (Git)Dokument98 SeitenPhysiology of The Gastrointestinal Tract (Git)jballungayNoch keine Bewertungen
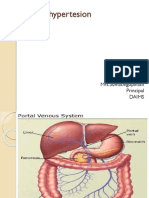
- Portal HypertensionDokument23 SeitenPortal HypertensionSumathi GopinathNoch keine Bewertungen
- Abdominal TraumaDokument30 SeitenAbdominal TraumaKavya KesavanNoch keine Bewertungen
- Animal Tissue - 2: Location FunctionDokument3 SeitenAnimal Tissue - 2: Location FunctionKb AshishNoch keine Bewertungen
- The Respiratory System: SubtitleDokument89 SeitenThe Respiratory System: SubtitleAaaNoch keine Bewertungen
- Anatomy of Bulbus VestibuliDokument6 SeitenAnatomy of Bulbus VestibuliSyahla FaizashaNoch keine Bewertungen
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Botany QuizDokument3 SeitenBotany QuizAleczandra QuesadaNoch keine Bewertungen
- CarcinomarectumDokument45 SeitenCarcinomarectumHuzefa MemonNoch keine Bewertungen
- The Respiratory SystemDokument6 SeitenThe Respiratory SystemIftikharNoch keine Bewertungen
- Respiratory and Circulatory Systems QuizDokument4 SeitenRespiratory and Circulatory Systems QuizGusthe Jebb AzarconNoch keine Bewertungen
- Maryam PresentationDokument9 SeitenMaryam PresentationUsman MuneerNoch keine Bewertungen
- Microflora PericoronariteDokument45 SeitenMicroflora PericoronariteCeresău MarinaNoch keine Bewertungen
- 02 Anatomy and Histology PLE 2019 RatioDokument69 Seiten02 Anatomy and Histology PLE 2019 RatioPatricia VillegasNoch keine Bewertungen
- Dental Management of Patients Under Anticoagulant TherapyDokument15 SeitenDental Management of Patients Under Anticoagulant TherapyThaer ZabenNoch keine Bewertungen
- Teeth Number 85Dokument16 SeitenTeeth Number 85rarasNoch keine Bewertungen
- Physiology lab: Faculty of Medical laboratory since Patch 7 ليعامسإ دمحم اللهدبع مكوخأ صيخلتDokument32 SeitenPhysiology lab: Faculty of Medical laboratory since Patch 7 ليعامسإ دمحم اللهدبع مكوخأ صيخلتmohammedNoch keine Bewertungen
- Abdominal Compartment SyndromeDokument12 SeitenAbdominal Compartment SyndromeAltus GoldenNoch keine Bewertungen
- General Biology1 (Answer Key)Dokument15 SeitenGeneral Biology1 (Answer Key)Olive PelayoNoch keine Bewertungen
- Angiosperm Reproduction and Biotechnology: BiologyDokument41 SeitenAngiosperm Reproduction and Biotechnology: BiologySem GacomsNoch keine Bewertungen
- Syllabus Breakup - Class 10 (Biology)Dokument2 SeitenSyllabus Breakup - Class 10 (Biology)AaraNoch keine Bewertungen
- Case StudyDokument41 SeitenCase StudychaSeph292784100% (6)
- DLP - ScienceDokument7 SeitenDLP - ScienceJUDE ABBUNoch keine Bewertungen
- Endocrine Disorders: Pathophysiology 2 0 2 1Dokument62 SeitenEndocrine Disorders: Pathophysiology 2 0 2 1Sawyer SprungNoch keine Bewertungen
- KayleeDokument3 SeitenKayleeKaylee RamsaroopNoch keine Bewertungen
- 1 TISSUES Online Teaching - REVISED 2021 AnaphyDokument38 Seiten1 TISSUES Online Teaching - REVISED 2021 AnaphySerendipity ParkNoch keine Bewertungen
- Clotting Factors List and Mnemonics For Easy Learning - All Medical StuffDokument10 SeitenClotting Factors List and Mnemonics For Easy Learning - All Medical Stuffmonika ramosNoch keine Bewertungen
- Crypt: Life, Death and Disease in the Middle Ages and BeyondVon EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondBewertung: 4 von 5 Sternen4/5 (3)
- Mitochondria and the Future of Medicine: The Key to Understanding Disease, Chronic Illness, Aging, and Life ItselfVon EverandMitochondria and the Future of Medicine: The Key to Understanding Disease, Chronic Illness, Aging, and Life ItselfBewertung: 4.5 von 5 Sternen4.5/5 (98)
- Why We Die: The New Science of Aging and the Quest for ImmortalityVon EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityBewertung: 3.5 von 5 Sternen3.5/5 (2)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyVon EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyBewertung: 3.5 von 5 Sternen3.5/5 (31)
- The Mind & The Brain: Neuroplasticity and the Power of Mental ForceVon EverandThe Mind & The Brain: Neuroplasticity and the Power of Mental ForceNoch keine Bewertungen
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessVon Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessBewertung: 4 von 5 Sternen4/5 (33)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisVon EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisBewertung: 3.5 von 5 Sternen3.5/5 (2)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldVon EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldBewertung: 4.5 von 5 Sternen4.5/5 (18)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesVon EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesBewertung: 4.5 von 5 Sternen4.5/5 (396)