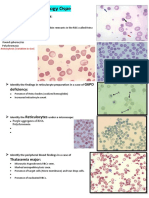
Das könnte Ihnen auch gefallen
- Post Exposure Prophylaxis For HivDokument21 SeitenPost Exposure Prophylaxis For Hivdr.a.k.gupta6924Noch keine Bewertungen
- Action Potential of Nerve FibreDokument2 SeitenAction Potential of Nerve Fibredr.a.k.gupta6924Noch keine Bewertungen
- Status of Orphan Vulnerable Children Infected or / Affected by HIV/AIDS in DelhiDokument32 SeitenStatus of Orphan Vulnerable Children Infected or / Affected by HIV/AIDS in Delhidr.a.k.gupta6924Noch keine Bewertungen
- Tracts of The Spinal CordDokument21 SeitenTracts of The Spinal Corddr.a.k.gupta6924100% (10)
- ElectrophoresisDokument2 SeitenElectrophoresisdr.a.k.gupta6924Noch keine Bewertungen
- Blood Physiology Part IV - Plasma Proteins, Hematocrit, ESR by DR A K GuptaDokument26 SeitenBlood Physiology Part IV - Plasma Proteins, Hematocrit, ESR by DR A K Guptadr.a.k.gupta6924Noch keine Bewertungen
- Physiology of Plasma ProteinsDokument21 SeitenPhysiology of Plasma Proteinsdr.a.k.gupta6924100% (4)
- HIV Intervention in College Youth by Red Ribbon ClubsDokument13 SeitenHIV Intervention in College Youth by Red Ribbon Clubsdr.a.k.gupta6924100% (1)
- GIT Physiology Part II by DR A K GutaDokument11 SeitenGIT Physiology Part II by DR A K Gutadr.a.k.gupta6924Noch keine Bewertungen
- Git Physiology Part IiiDokument9 SeitenGit Physiology Part Iiidr.a.k.gupta6924100% (1)
- Holistic Care of People Living With HIV/AIDS by DR A.K. Gupta, DSACSDokument36 SeitenHolistic Care of People Living With HIV/AIDS by DR A.K. Gupta, DSACSdr.a.k.gupta6924100% (1)
- Post Exposure Prophylaxis & Universal Work Precautions - PPDokument43 SeitenPost Exposure Prophylaxis & Universal Work Precautions - PPdr.a.k.gupta6924Noch keine Bewertungen
- Natural History of HIV Infection by DR A K Gupta, Addl. Project Director, DSACSDokument29 SeitenNatural History of HIV Infection by DR A K Gupta, Addl. Project Director, DSACSdr.a.k.gupta6924100% (3)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- II.a. Cell The Basic Unit of LifeDokument27 SeitenII.a. Cell The Basic Unit of LifeDarwin NoolNoch keine Bewertungen
- 1Dokument12 Seiten1ShuvashishSunuwarNoch keine Bewertungen
- Bem-Aventurados Os Simples (Psicografia Waldo Vieira - Espírito Valérium)Dokument673 SeitenBem-Aventurados Os Simples (Psicografia Waldo Vieira - Espírito Valérium)Alexsandro ClaudinoNoch keine Bewertungen
- Hematology Ospe A1Dokument5 SeitenHematology Ospe A1Zain Ul AbidinNoch keine Bewertungen
- Cell NucleusDokument17 SeitenCell NucleusAsad Aly100% (8)
- Cell Biology NotesDokument6 SeitenCell Biology Notessaadc786100% (1)
- EBIO-Table On Chromosomal Abnormalities-1Dokument2 SeitenEBIO-Table On Chromosomal Abnormalities-1Alyanna Elisse VergaraNoch keine Bewertungen
- By DR Ananya MandalDokument4 SeitenBy DR Ananya MandalhasimNoch keine Bewertungen
- Karyotype Activity GoodDokument12 SeitenKaryotype Activity GoodSATZU MALAKASNoch keine Bewertungen
- Anatomy and Physiology DefinedDokument78 SeitenAnatomy and Physiology DefinedFhen Farrel100% (5)
- 5th Sem Syllabus BotanyDokument3 Seiten5th Sem Syllabus BotanyAdnan BandayNoch keine Bewertungen
- Class 6 ScienceDokument8 SeitenClass 6 Sciencemuzzammil.memonsNoch keine Bewertungen
- Oranur 1Dokument4 SeitenOranur 1abyssarmyNoch keine Bewertungen
- Parts of A Typical Cell and Their FunctionsDokument2 SeitenParts of A Typical Cell and Their FunctionsMaria Sahara FregilNoch keine Bewertungen
- Research Article: Involvement of Prohibitin Upregulation in Abrin-Triggered ApoptosisDokument11 SeitenResearch Article: Involvement of Prohibitin Upregulation in Abrin-Triggered ApoptosisAndri Praja SatriaNoch keine Bewertungen
- Cell CycleDokument69 SeitenCell CycleJustine FariscalNoch keine Bewertungen
- MT1 HSCI LEC L7 HematopoiesisDokument10 SeitenMT1 HSCI LEC L7 HematopoiesisSEBASTIEN ANDREI BUENAFENoch keine Bewertungen
- 8th Biology English PDFDokument167 Seiten8th Biology English PDFVadlakonda SampathgoudNoch keine Bewertungen
- Vocabulary CellsDokument6 SeitenVocabulary CellsJorge LunaNoch keine Bewertungen
- Biochemistry Free For All FinalDokument3.578 SeitenBiochemistry Free For All Finalharsh100% (1)
- Cell Structure & FunctionDokument38 SeitenCell Structure & Functiongundogan21100% (1)
- Basic Concepts in MicrobiologyDokument4 SeitenBasic Concepts in MicrobiologyHazelle RoxasNoch keine Bewertungen
- Lecture - 1: Nafis Neehal, Lecturer, Department of CSE, DIUDokument21 SeitenLecture - 1: Nafis Neehal, Lecturer, Department of CSE, DIUsk3 khanNoch keine Bewertungen
- AnaPhy-lab ACTSDokument5 SeitenAnaPhy-lab ACTSCEEJNoch keine Bewertungen
- Improvisation of Conventional Techniques: The Future of Oncology ResearchDokument3 SeitenImprovisation of Conventional Techniques: The Future of Oncology ResearchasclepiuspdfsNoch keine Bewertungen
- CellDokument33 SeitenCellbrij100% (1)
- Transcription: RNA Polymerases and General Transcription FactorsDokument77 SeitenTranscription: RNA Polymerases and General Transcription FactorsmluluNoch keine Bewertungen
- 21b - Cell - Structure - Function 2 &quizDokument54 Seiten21b - Cell - Structure - Function 2 &quizammy adeNoch keine Bewertungen
- Class 9 - Cell - Structural and Functional Unit of LifeDokument40 SeitenClass 9 - Cell - Structural and Functional Unit of LifeanyastudysNoch keine Bewertungen
- Cells and Cell StructureDokument3 SeitenCells and Cell Structureapi-238197463Noch keine Bewertungen