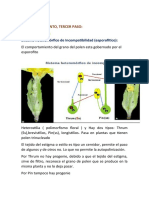
Das könnte Ihnen auch gefallen
- 7 Poliploidia 2011Dokument38 Seiten7 Poliploidia 2011Daisy VanessaNoch keine Bewertungen
- 14 HaploidiaPoliploidiaDokument85 Seiten14 HaploidiaPoliploidiaLeon PaineNoch keine Bewertungen
- Tema 9 PoliploidiaDokument30 SeitenTema 9 PoliploidiaAdolf MarNoch keine Bewertungen
- La Poliploidia en El FitomejoramientoDokument15 SeitenLa Poliploidia en El FitomejoramientoSteice Gabriela Lescano OlivaNoch keine Bewertungen
- Poli Plo IdesDokument30 SeitenPoli Plo IdesCristhian Garcia Vilchez100% (1)
- Poliploidia V 2023-2Dokument35 SeitenPoliploidia V 2023-2PABLO ELIAS PEREZ CANELANoch keine Bewertungen
- Deber 3 Genetica Agricola - 2do ParcialDokument11 SeitenDeber 3 Genetica Agricola - 2do Parcialkevin acuñaNoch keine Bewertungen
- 6ta Semana 6ta Clase TeoriaDokument26 Seiten6ta Semana 6ta Clase TeoriaJair RamosNoch keine Bewertungen
- Tema 10 PoliploidiaDokument30 SeitenTema 10 PoliploidiaDanielAnthonyIngaAnccoNoch keine Bewertungen
- (002805) PDFDokument99 Seiten(002805) PDFgraulNoch keine Bewertungen
- Mutacion GeneticaDokument43 SeitenMutacion GeneticaWilson Gonzalo T QNoch keine Bewertungen
- CLASE 9. Mutación y Poliploidia PDFDokument35 SeitenCLASE 9. Mutación y Poliploidia PDFYassmin Huayta FierroNoch keine Bewertungen
- Clase 12. Plantas PoliploidesDokument20 SeitenClase 12. Plantas Poliploidesmeliodas ruokNoch keine Bewertungen
- 28-03-2020 Unidad I y IV Mejoramiento en FrutalesDokument32 Seiten28-03-2020 Unidad I y IV Mejoramiento en FrutalesManuel Ulises VarelaNoch keine Bewertungen
- DIPLOIDEDokument5 SeitenDIPLOIDEGEYSLER MARIÑAS ALVARADONoch keine Bewertungen
- Resumen - CitogeneticaDokument3 SeitenResumen - CitogeneticaAngela DávilaNoch keine Bewertungen
- Genetica y Determinación de Los Grupos Sanguíneos y Del Factor RHDokument12 SeitenGenetica y Determinación de Los Grupos Sanguíneos y Del Factor RHLucero Odar CastilloNoch keine Bewertungen
- POLIPLOIDEDokument9 SeitenPOLIPLOIDELuna IrfanNoch keine Bewertungen
- Citogenética II.23Dokument25 SeitenCitogenética II.23victorcondor666Noch keine Bewertungen
- Qué Es La PoliploidíaDokument6 SeitenQué Es La Poliploidíaesneider sanchezNoch keine Bewertungen
- Biotecnologia y Mejoramiento GeneticoDokument91 SeitenBiotecnologia y Mejoramiento GeneticoHer R MinattiNoch keine Bewertungen
- CITOCINESISDokument3 SeitenCITOCINESISLuis Carlos Barraza CarvalNoch keine Bewertungen
- Mutaciones y PoliploidiaDokument9 SeitenMutaciones y PoliploidiaMelissa HesterNoch keine Bewertungen
- Tercer Paso de FitomejoDokument10 SeitenTercer Paso de FitomejoyulizzaNoch keine Bewertungen
- PoliploidiaDokument4 SeitenPoliploidiaLalo TolentinoNoch keine Bewertungen
- Genoma de La PapaDokument1 SeiteGenoma de La PapaSarayBurbanoNoch keine Bewertungen
- PloidíaDokument2 SeitenPloidíaLiliana RivasNoch keine Bewertungen
- Poliploidia Principal Rol Fitomejoramiento 2016Dokument18 SeitenPoliploidia Principal Rol Fitomejoramiento 2016igarashigantadeadmanwonderlandNoch keine Bewertungen
- Mejoramiento - YucaDokument47 SeitenMejoramiento - YucaGroverNoch keine Bewertungen
- Consulta HaploideDokument4 SeitenConsulta HaploideAnniee FelizNoch keine Bewertungen
- Variaciones CromosomicasDokument4 SeitenVariaciones CromosomicasHugo CartonNoch keine Bewertungen
- Mitosis Meiosis Definición ¿Ocurre en Todos Los Tejidos? Reproducción Sexual Reproducción AsexualDokument10 SeitenMitosis Meiosis Definición ¿Ocurre en Todos Los Tejidos? Reproducción Sexual Reproducción AsexualLeidi JNoch keine Bewertungen
- Variaciones CromosomicasDokument35 SeitenVariaciones CromosomicasYhasmira Rojas Aguirre100% (5)
- Estructura Genetica de Poblaciones 2012Dokument36 SeitenEstructura Genetica de Poblaciones 2012Erick MendezNoch keine Bewertungen
- Variabilidad GenéticaDokument54 SeitenVariabilidad Genéticadiana_hernandez_88Noch keine Bewertungen
- Poliploidia e Hibridacion Interespecifica 2011 para ColgarDokument35 SeitenPoliploidia e Hibridacion Interespecifica 2011 para ColgarNestor Omar AmbesiNoch keine Bewertungen
- Variaciones Cromosomicas NumericasDokument99 SeitenVariaciones Cromosomicas NumericasJohnny DelvaNoch keine Bewertungen
- Poliploidia e Hibridacion Interespecifica PDFDokument28 SeitenPoliploidia e Hibridacion Interespecifica PDFJhonel Trujillo Ortiz100% (1)
- Herencia y GenéticaDokument1 SeiteHerencia y GenéticaArtic ANoch keine Bewertungen
- Trabajo de Biologia MaraDokument18 SeitenTrabajo de Biologia MaraMara VacaNoch keine Bewertungen
- Importancia de La Papa en El PerúDokument40 SeitenImportancia de La Papa en El PerúCorina Lucero IGNoch keine Bewertungen
- 03 - Dihaploids - En.es TRADUCIDODokument29 Seiten03 - Dihaploids - En.es TRADUCIDOolga avilaNoch keine Bewertungen
- TALLER 7 GeneticaDokument9 SeitenTALLER 7 GeneticaHector Estrada MarinNoch keine Bewertungen
- Herencia CitoplasmicaDokument15 SeitenHerencia CitoplasmicaIvoshka Ramos BecerraNoch keine Bewertungen
- Orquideas Duplicacion Cromosomas 24-28Dokument5 SeitenOrquideas Duplicacion Cromosomas 24-28Lina marcela Cano VillarragaNoch keine Bewertungen
- Clase N - 8 Mejoramiento de Plantas AutogamasDokument160 SeitenClase N - 8 Mejoramiento de Plantas AutogamasJeancarlo Moori50% (4)
- Genetic PloidiasDokument16 SeitenGenetic PloidiasObi ChavezNoch keine Bewertungen
- Citogenética PARA ESTUDIANTESDokument51 SeitenCitogenética PARA ESTUDIANTESdonovan ojitoNoch keine Bewertungen
- Taller Gnética MendelianaDokument8 SeitenTaller Gnética MendelianavalentinaNoch keine Bewertungen
- Investigacion Androesterilidad FitomejoramientoDokument14 SeitenInvestigacion Androesterilidad Fitomejoramientoedwininteriano49Noch keine Bewertungen
- Mejoramiento Genético (Métodos)Dokument2 SeitenMejoramiento Genético (Métodos)Alex Valle SotoNoch keine Bewertungen
- Teoria Reproduccion Sexual PDFDokument10 SeitenTeoria Reproduccion Sexual PDFgatinha33Noch keine Bewertungen
- TP Nº11Dokument3 SeitenTP Nº11Maria Florencia MelliaNoch keine Bewertungen
- Unidad 01 Bases Fundamentales Del FitomejoramientoDokument58 SeitenUnidad 01 Bases Fundamentales Del FitomejoramientoVivian ArdónNoch keine Bewertungen
- 1er-Examen Fitomejoramiento (Completo)Dokument5 Seiten1er-Examen Fitomejoramiento (Completo)Diego Torres SarzozoNoch keine Bewertungen
- IV Bim - 5to. Año - Bio - Guía 2 - Genética IDokument4 SeitenIV Bim - 5to. Año - Bio - Guía 2 - Genética IAngel Stevens Garcia CallaoNoch keine Bewertungen
- Informe Biologia GeneticaDokument20 SeitenInforme Biologia GeneticaDarío LópezNoch keine Bewertungen
- Gms12 Jara RaquelDokument15 SeitenGms12 Jara Raquel7z6v7d7dhyNoch keine Bewertungen
- ESTERILIDADDokument8 SeitenESTERILIDADhopeNoch keine Bewertungen
- Lista Anotada de Los Peces de Aguas Continentales Del Perú. 2012Dokument59 SeitenLista Anotada de Los Peces de Aguas Continentales Del Perú. 2012Roger Barboza CastroNoch keine Bewertungen
- Diversidad de Mamiferos de Los Bosques M PDFDokument20 SeitenDiversidad de Mamiferos de Los Bosques M PDFRoger Barboza CastroNoch keine Bewertungen
- Expediente Tecnico ACP Bosque de CuyasDokument26 SeitenExpediente Tecnico ACP Bosque de CuyasRoger Barboza Castro100% (1)
- Tesis - Ornitofauna Otuzco. Alegre. 2016Dokument75 SeitenTesis - Ornitofauna Otuzco. Alegre. 2016Roger Barboza CastroNoch keine Bewertungen
- Polylepis EcologiaDokument99 SeitenPolylepis EcologiaRoger Barboza CastroNoch keine Bewertungen
- Métodos de Observacion en EtologíaDokument5 SeitenMétodos de Observacion en EtologíaRoger Barboza CastroNoch keine Bewertungen
- Gral Buffer TAE TBEDokument2 SeitenGral Buffer TAE TBERoger Barboza Castro50% (2)
- Atlas de Los Escarabajos de Mexico, Scarabaeidae, Trogidae, Passalidae y LucanidaeDokument224 SeitenAtlas de Los Escarabajos de Mexico, Scarabaeidae, Trogidae, Passalidae y LucanidaePablo Ramos GaleanaNoch keine Bewertungen
- 2) Anexo 7. Cal. Censo For (Nuevo Formato Marzo 2023) - E.globulusDokument2 Seiten2) Anexo 7. Cal. Censo For (Nuevo Formato Marzo 2023) - E.globulusDavid Arrobo GarcíaNoch keine Bewertungen
- Obtención Del Aceite Esencial Por Arrastre de VaporDokument12 SeitenObtención Del Aceite Esencial Por Arrastre de VaporgreciasaonaNoch keine Bewertungen
- Forraje y Clasificacion (Sem 2)Dokument24 SeitenForraje y Clasificacion (Sem 2)popi50% (2)
- Diferencia Entre El Café Arábica y El Café RobustaDokument3 SeitenDiferencia Entre El Café Arábica y El Café RobustaRamiro OrtizNoch keine Bewertungen
- Laboratorio Estructura y Diversidad CelularDokument19 SeitenLaboratorio Estructura y Diversidad CelularAnderson HernandezNoch keine Bewertungen
- 1..encuesta Terminada Pa PresentarDokument40 Seiten1..encuesta Terminada Pa PresentarMichael BrenisNoch keine Bewertungen
- FitomejoramientoDokument10 SeitenFitomejoramientoMario Andres LassoNoch keine Bewertungen
- Informe de PracticasDokument83 SeitenInforme de PracticasMary Torres VasquezNoch keine Bewertungen
- Alta Tension Reglamento (Es)Dokument12 SeitenAlta Tension Reglamento (Es)josepNoch keine Bewertungen
- Clasificacion de FrutalesDokument3 SeitenClasificacion de FrutalesYuder M CurahuaNoch keine Bewertungen
- Alimentación de Gallinas y PatosDokument4 SeitenAlimentación de Gallinas y Patosgtss35Noch keine Bewertungen
- Seminario Análisis CombinatorioDokument2 SeitenSeminario Análisis Combinatoriocamila hernandezNoch keine Bewertungen
- Aceite de LimonDokument3 SeitenAceite de LimonCristian CCNoch keine Bewertungen
- Palma de AceiteDokument3 SeitenPalma de AceiteEliana Patricia Negrete GomezNoch keine Bewertungen
- La ManzanillaDokument5 SeitenLa ManzanillaesthefaniNoch keine Bewertungen
- Plagas Cuarentenarias en Citricos Importancia y Avances eDokument73 SeitenPlagas Cuarentenarias en Citricos Importancia y Avances eSantiago MartínezNoch keine Bewertungen
- 2021CIS708 FirmadoDokument5 Seiten2021CIS708 FirmadoreiferNoch keine Bewertungen
- Clasificacion Ecologica de BoliviaDokument35 SeitenClasificacion Ecologica de BoliviaBasilioQuispeDonaireNoch keine Bewertungen
- Presentación Aceites EsencialesDokument7 SeitenPresentación Aceites EsencialesMaria BustamanteNoch keine Bewertungen
- Danzas, Trajes, Comida, Etc..de GuatemalaDokument22 SeitenDanzas, Trajes, Comida, Etc..de GuatemalaHeidi Diana StaeheliNoch keine Bewertungen
- TRIPTICODokument2 SeitenTRIPTICOBayrin Hernandez100% (1)
- Alimentacion SaludableDokument34 SeitenAlimentacion SaludableVivianOspinaClavijoNoch keine Bewertungen
- Lista Actual Del Personal Del Proyecto 2023Dokument7 SeitenLista Actual Del Personal Del Proyecto 2023Luis Mamani MaydanaNoch keine Bewertungen
- Vegetacion La RiojaDokument32 SeitenVegetacion La RiojadigitalNoch keine Bewertungen
- Mecanica de Fluidos - Trabajo Practico - Huancahuire Paredes SebastianDokument19 SeitenMecanica de Fluidos - Trabajo Practico - Huancahuire Paredes Sebastianchrystin2009Noch keine Bewertungen
- Reforestacion en Manglares de Oaxaca PDFDokument65 SeitenReforestacion en Manglares de Oaxaca PDFAndreaNoch keine Bewertungen
- D11T Tractor de Cadenas Sistema HidráulicoDokument7 SeitenD11T Tractor de Cadenas Sistema HidráulicoJulio Edgar QuiritaNoch keine Bewertungen
- Comentarios, Correcciones y Anotaciones Sobre Pompílidos (Hymenoptera Pompilidae) en Algunos Libros Chilenos.Dokument8 SeitenComentarios, Correcciones y Anotaciones Sobre Pompílidos (Hymenoptera Pompilidae) en Algunos Libros Chilenos.Víctor Antonio Broncano MallquiNoch keine Bewertungen
- VI Simposio Internacional de Biotecnologia VegetalDokument154 SeitenVI Simposio Internacional de Biotecnologia VegetalMiriamLopezVazquezNoch keine Bewertungen