Das könnte Ihnen auch gefallen
- Aneesb PDFDokument26 SeitenAneesb PDFSufail ANoch keine Bewertungen
- Health Promotion and Disease PreventionDokument21 SeitenHealth Promotion and Disease PreventionHajra MirzaNoch keine Bewertungen
- Head To Toe Assessment WorksheetDokument7 SeitenHead To Toe Assessment WorksheetNina Klairea PaduaNoch keine Bewertungen
- Surgery Question From AbeideDokument44 SeitenSurgery Question From Abeide69xXALModr3EMXx69 McAnusNoch keine Bewertungen
- Primary Health CareDokument7 SeitenPrimary Health CareJesena SalveNoch keine Bewertungen
- Splinting Techniques: by Madhumitha M, Aruna G, Abi P, Sakthivelayudham G, Sujidha R, Srivignesh MDokument99 SeitenSplinting Techniques: by Madhumitha M, Aruna G, Abi P, Sakthivelayudham G, Sujidha R, Srivignesh MSakthivelayudhamganesanNoch keine Bewertungen
- The Human Metabolic Response To Chronic Ketosis Without Caloric Restriction - 1Dokument12 SeitenThe Human Metabolic Response To Chronic Ketosis Without Caloric Restriction - 1acolpoNoch keine Bewertungen
- SEM-11-Starvation Saudek Felig 1976Dokument10 SeitenSEM-11-Starvation Saudek Felig 1976TomasNoch keine Bewertungen
- CelluloseDokument7 SeitenCellulosem waltersNoch keine Bewertungen
- 3.4 Paralytic Ileus Due To Potassium Depletion. Dr. S. GieveDokument3 Seiten3.4 Paralytic Ileus Due To Potassium Depletion. Dr. S. GieveDaru KristiyonoNoch keine Bewertungen
- J. Biol. Chem.-1918-Harding-575-80Dokument7 SeitenJ. Biol. Chem.-1918-Harding-575-80Vincenzo Paolo FraddosioNoch keine Bewertungen
- Nutritional and Protein: Albumin Metabolism: Effect of The State The IntakeDokument15 SeitenNutritional and Protein: Albumin Metabolism: Effect of The State The IntakeDio PattersonNoch keine Bewertungen
- Role of Sugars in Human Neutrophilic PhagocytosisDokument5 SeitenRole of Sugars in Human Neutrophilic Phagocytosisdumitrudragos100% (2)
- Factors Affecting Gluconeogenesis in The Neonatal Subhuman Primate (Macaca Mulatta)Dokument8 SeitenFactors Affecting Gluconeogenesis in The Neonatal Subhuman Primate (Macaca Mulatta)Saleth CortezNoch keine Bewertungen
- Spontaneous 24-h Ghrelin Secretion Pattern in FastDokument7 SeitenSpontaneous 24-h Ghrelin Secretion Pattern in Fastmv.Noch keine Bewertungen
- Evaluation of Anti-Aging Effects of Gemfibrozil OnDokument6 SeitenEvaluation of Anti-Aging Effects of Gemfibrozil OnMario CoelhoNoch keine Bewertungen
- Extraction of Glycogen On Mild Condition Lacks Aig FractionDokument7 SeitenExtraction of Glycogen On Mild Condition Lacks Aig FractionMaika DoronelaNoch keine Bewertungen
- Philippe Ducrotté and Pierre Déchelotte Moïse Coëffier, Sophie Claeyssens, Bernadette Hecketsweiler, Alain LavoinneDokument9 SeitenPhilippe Ducrotté and Pierre Déchelotte Moïse Coëffier, Sophie Claeyssens, Bernadette Hecketsweiler, Alain LavoinneMoonsferatuNoch keine Bewertungen
- Clinical Nutrition: Original ArticleDokument6 SeitenClinical Nutrition: Original ArticleS Dian RNoch keine Bewertungen
- 8 HalamansdbDokument14 Seiten8 HalamansdbbanglarkNoch keine Bewertungen
- ImplantDokument8 SeitenImplantTabishur RahmanNoch keine Bewertungen
- 2019 RMC1Dokument16 Seiten2019 RMC1corinaNoch keine Bewertungen
- J. Lipid Res.-2007-Varady-2212-9Dokument8 SeitenJ. Lipid Res.-2007-Varady-2212-9ggutierrezdieckNoch keine Bewertungen
- Vol 2 Iss 2 Page 1-4 Demonstrating Glycogen at Various Stages of Developing Human LiverDokument4 SeitenVol 2 Iss 2 Page 1-4 Demonstrating Glycogen at Various Stages of Developing Human LiverEditorsNoch keine Bewertungen
- Lead Intoxication in A Puppy - A Case Report: E. S, J. P - G, I. K, B. P, B. A, Z. Š - PDokument2 SeitenLead Intoxication in A Puppy - A Case Report: E. S, J. P - G, I. K, B. P, B. A, Z. Š - PMauricio RíosNoch keine Bewertungen
- Jurnal PK FaridaDokument34 SeitenJurnal PK FaridaAdelia Dwi KusumaNoch keine Bewertungen
- Estrogen Prevents Increased Hepatic Aquaporin-9 Expression and Glycerol Uptake During StarvationDokument10 SeitenEstrogen Prevents Increased Hepatic Aquaporin-9 Expression and Glycerol Uptake During StarvationMarta SandeNoch keine Bewertungen
- GLP AlzhmrDokument14 SeitenGLP Alzhmryengeç adamNoch keine Bewertungen
- Sustained Hyperhydration With Glycerol IngestionDokument9 SeitenSustained Hyperhydration With Glycerol IngestionBud Marvin LeRoy RiedeselNoch keine Bewertungen
- tmpD4AC TMPDokument14 SeitentmpD4AC TMPFrontiersNoch keine Bewertungen
- Economic Pigging Cycles For Low-Throughput PipelinesDokument11 SeitenEconomic Pigging Cycles For Low-Throughput Pipelineseng.Abdelaziz RedaNoch keine Bewertungen
- Extended Definition RevisedDokument6 SeitenExtended Definition Revisedapi-491301139Noch keine Bewertungen
- Intermittent Fasting and Refeeding On Insulin Dependant MenDokument10 SeitenIntermittent Fasting and Refeeding On Insulin Dependant MenTyrone PenningNoch keine Bewertungen
- Cuy BeditoDokument15 SeitenCuy BeditoLucero CarranzaNoch keine Bewertungen
- Hons2009 PDFDokument11 SeitenHons2009 PDFBkash ShahNoch keine Bewertungen
- Fasting REEDokument5 SeitenFasting REEAimone FerriNoch keine Bewertungen
- Глюкагон мен глюкоза, инсулинDokument5 SeitenГлюкагон мен глюкоза, инсулинArsen IzbanovNoch keine Bewertungen
- Renal Glycosuria Case StudyDokument2 SeitenRenal Glycosuria Case StudyRodeoKenan100% (1)
- Jcinvest00248 0177 PDFDokument11 SeitenJcinvest00248 0177 PDFluis morales garzaNoch keine Bewertungen
- 1472 6882 2 1 PDFDokument7 Seiten1472 6882 2 1 PDFThiago NunesNoch keine Bewertungen
- Antidiabetic and Long-Term Effects of Elaeocarpus GrandiflorusDokument12 SeitenAntidiabetic and Long-Term Effects of Elaeocarpus GrandiflorusAgus FakhrudinNoch keine Bewertungen
- Effect of Intermittent Fasting and Refeeding On Insulin Action in Healthy MenDokument9 SeitenEffect of Intermittent Fasting and Refeeding On Insulin Action in Healthy Mendgh3Noch keine Bewertungen
- Response of C57Bl/6 Mice To A Carbohydrate-Free Diet: Research Open AccessDokument6 SeitenResponse of C57Bl/6 Mice To A Carbohydrate-Free Diet: Research Open AccesspopopioNoch keine Bewertungen
- Colonie Function and Fermentation in Men Consuming High Fiber Diets1Dokument10 SeitenColonie Function and Fermentation in Men Consuming High Fiber Diets1MuzayyanahNoch keine Bewertungen
- Chlorogenic Acid and Caffeic Acid Are Absorbed in HumansDokument6 SeitenChlorogenic Acid and Caffeic Acid Are Absorbed in HumansluisbrumovskyNoch keine Bewertungen
- Edr2010 289614Dokument10 SeitenEdr2010 289614FawwaziNoch keine Bewertungen
- Thiopropanol Induced Changes in Glycogen Breakdown in Alloxan Diabetic LiverDokument5 SeitenThiopropanol Induced Changes in Glycogen Breakdown in Alloxan Diabetic LiverNirmaLa SariNoch keine Bewertungen
- Brief Report Water Drinking Induces Thermogenesis Through Osmosensitive MechanismsDokument4 SeitenBrief Report Water Drinking Induces Thermogenesis Through Osmosensitive MechanismsS Dian RNoch keine Bewertungen
- Effects of Starvation On Proximate Chemical Composition and Energy Utilization of Plaice, LDokument10 SeitenEffects of Starvation On Proximate Chemical Composition and Energy Utilization of Plaice, Ldaniel cretuNoch keine Bewertungen
- Obesity - 2012 - Sampey - Cafeteria Diet Is A Robust Model of Human Metabolic Syndrome With Liver and Adipose InflammationDokument10 SeitenObesity - 2012 - Sampey - Cafeteria Diet Is A Robust Model of Human Metabolic Syndrome With Liver and Adipose InflammationRodrigoNoch keine Bewertungen
- MD 1 Section E2 Group 3 Biochem REVISEDDokument10 SeitenMD 1 Section E2 Group 3 Biochem REVISEDParadoxistNoch keine Bewertungen
- WAT Plasticity in Response To Beta3-ARDokument9 SeitenWAT Plasticity in Response To Beta3-ARassaf.rudichNoch keine Bewertungen
- Efecto de CetonaDokument9 SeitenEfecto de CetonaGiusi CaforioNoch keine Bewertungen
- Mechanisms of Sleep Deprivation-Induced Hepatic Steatosis and Insulin Resistance in MiceDokument11 SeitenMechanisms of Sleep Deprivation-Induced Hepatic Steatosis and Insulin Resistance in MiceNazan ElmaNoch keine Bewertungen
- Efectividad de Colageno Hidrolizado CientificoDokument9 SeitenEfectividad de Colageno Hidrolizado CientificoRojas Evert AlonsoNoch keine Bewertungen
- The Clinical, And: Nephrotic Syndrome of Childhood: Immunologic, Pathologic CorrelationsDokument11 SeitenThe Clinical, And: Nephrotic Syndrome of Childhood: Immunologic, Pathologic Correlationsrizal_yahya_1Noch keine Bewertungen
- Ginger PublishedDokument8 SeitenGinger PublishedFatimoh AbdulsalamNoch keine Bewertungen
- Exp. 6: Isolation of GlycogenDokument7 SeitenExp. 6: Isolation of GlycogenBelaNoch keine Bewertungen
- tmp2FD1 TMPDokument11 Seitentmp2FD1 TMPFrontiersNoch keine Bewertungen
- Resveratrol Attenuates Oxidative Stress and Histological Alterations Induced by Liver Ischemia/reperfusion in RatsDokument6 SeitenResveratrol Attenuates Oxidative Stress and Histological Alterations Induced by Liver Ischemia/reperfusion in RatsAbi ZakiNoch keine Bewertungen
- Javma 237 3 299Dokument5 SeitenJavma 237 3 299tcvhNoch keine Bewertungen
- Jurnal InternationalDokument9 SeitenJurnal InternationalAngelNoch keine Bewertungen
- Ketoacidosis in A Non-Diabetic Patient: Clinical VignetteDokument2 SeitenKetoacidosis in A Non-Diabetic Patient: Clinical VignetteDr ZapataNoch keine Bewertungen
- Case StudyDokument3 SeitenCase StudyAnnie Laiza BacayNoch keine Bewertungen
- European Journal of Obstetrics & Gynecology and Reproductive BiologyDokument8 SeitenEuropean Journal of Obstetrics & Gynecology and Reproductive BiologyJose Alejandro InciongNoch keine Bewertungen
- Pathogens and Immune Response of CephalopodsDokument9 SeitenPathogens and Immune Response of CephalopodsJose Alejandro InciongNoch keine Bewertungen
- Compound Astragalus and Salvia Miltiorrhiza Extracts Modulate MAPK Regulated TGF Smad Signaling in Hepatocellular Carcinoma by Multi Target MechanismDokument10 SeitenCompound Astragalus and Salvia Miltiorrhiza Extracts Modulate MAPK Regulated TGF Smad Signaling in Hepatocellular Carcinoma by Multi Target MechanismJose Alejandro InciongNoch keine Bewertungen
- Isolation and Characterization of Novel Protein With Anti-Fungal and Anti-Inflammatory Properties From Aloe Vera Leaf GelDokument7 SeitenIsolation and Characterization of Novel Protein With Anti-Fungal and Anti-Inflammatory Properties From Aloe Vera Leaf GelJose Alejandro Inciong100% (1)
- Anaplastic Sarcoma of The Kidney With Chromosomal AbnormalityDokument5 SeitenAnaplastic Sarcoma of The Kidney With Chromosomal AbnormalityJose Alejandro InciongNoch keine Bewertungen
- Ceohns Disease ECCO Guideline SumarizedDokument19 SeitenCeohns Disease ECCO Guideline Sumarizednishad nilangaNoch keine Bewertungen
- Final Patient Scenario Chapter 44 Nursing Care of Family When A Child Has A Hematologic DisorderDokument53 SeitenFinal Patient Scenario Chapter 44 Nursing Care of Family When A Child Has A Hematologic DisorderDay Meds100% (1)
- Summative Test No 4 in MapehDokument2 SeitenSummative Test No 4 in MapehDianaRoseAcupeadoNoch keine Bewertungen
- APHASIADokument131 SeitenAPHASIArnsharmasNoch keine Bewertungen
- Pathophysiology of Heart Failure With Reduced Ejection Fraction - Hemodynamic Alterations and Remodeling - UpToDateDokument29 SeitenPathophysiology of Heart Failure With Reduced Ejection Fraction - Hemodynamic Alterations and Remodeling - UpToDatesamNoch keine Bewertungen
- Sample Email For Leave Request To ManagerDokument19 SeitenSample Email For Leave Request To Managersaikiran eligetiNoch keine Bewertungen
- Case Study 3 CVD Bleed HCVD Type 2 DM CICD Stage 5 WorksheetDokument27 SeitenCase Study 3 CVD Bleed HCVD Type 2 DM CICD Stage 5 WorksheetJade HemmingsNoch keine Bewertungen
- Antimicrobial Activity of Citrus SinensisDokument44 SeitenAntimicrobial Activity of Citrus SinensisherrNoch keine Bewertungen
- Nails in Nutritional DefinciesDokument5 SeitenNails in Nutritional DefinciesstcerieNoch keine Bewertungen
- Pressure Injury - Nursing Care PlanDokument4 SeitenPressure Injury - Nursing Care PlanMonique Sacherow BacherNoch keine Bewertungen
- Medicinal Chemistry I Sovan Sarkar 186012111012Dokument11 SeitenMedicinal Chemistry I Sovan Sarkar 186012111012Sovan SarkarNoch keine Bewertungen
- Annex 1Dokument1 SeiteAnnex 1Shweh Fern LooNoch keine Bewertungen
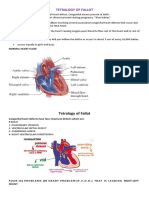
- Tetralogy of Fallot Report 1Dokument9 SeitenTetralogy of Fallot Report 1Maricar EustaquioNoch keine Bewertungen
- Project #1-Caregiver's BrochureDokument2 SeitenProject #1-Caregiver's BrochureswarnaNoch keine Bewertungen
- Prophylactic Antibiotics For HemorrhoidectomyDokument5 SeitenProphylactic Antibiotics For HemorrhoidectomyThắng NguyễnNoch keine Bewertungen
- Catalogue - DP100 (Inno) PDFDokument6 SeitenCatalogue - DP100 (Inno) PDFHaliun AltangerelNoch keine Bewertungen
- Growth Chart For Boys Birth To 36 MonthsDokument2 SeitenGrowth Chart For Boys Birth To 36 MonthsCarlos TejedaNoch keine Bewertungen
- YH Neonatal Paediatric Hyperammonaemia GuidelineDokument9 SeitenYH Neonatal Paediatric Hyperammonaemia GuidelineTrishenth FonsekaNoch keine Bewertungen
- Miano Et Al-2022-Intensive Care MedicineDokument12 SeitenMiano Et Al-2022-Intensive Care MedicinelamcvNoch keine Bewertungen
- Unit 1: A Long and Healthy Life: Bacteria Disease Energy Fitness Muscles Tuberculosis Viruses StrengthDokument4 SeitenUnit 1: A Long and Healthy Life: Bacteria Disease Energy Fitness Muscles Tuberculosis Viruses StrengthLinh LamNoch keine Bewertungen
- Fundamentals in Nursing TransDokument14 SeitenFundamentals in Nursing TransPia Gabrielle CabatanaNoch keine Bewertungen
- Paediatric Pharmacy Services Guideline PDFDokument213 SeitenPaediatric Pharmacy Services Guideline PDFFazilah Shahrom100% (1)
- EXAM B SolDokument10 SeitenEXAM B SolNuhad Bou MoslehNoch keine Bewertungen
- Fastep COVID-19 IgG-IgM 英文说明书(黑白A4双面双胶)2页Dokument2 SeitenFastep COVID-19 IgG-IgM 英文说明书(黑白A4双面双胶)2页Nia AmbarwatiNoch keine Bewertungen