Das könnte Ihnen auch gefallen
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (119)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (587)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2219)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (894)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Social desirability bias and Freud's theories of the unconscious mindDokument13 SeitenSocial desirability bias and Freud's theories of the unconscious mindSay77% (13)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- Its Only A PenisDokument29 SeitenIts Only A PenisAdam BlairNoch keine Bewertungen
- Effect of Gender On Short Term Memory FinishedDokument27 SeitenEffect of Gender On Short Term Memory Finishedhafsah286Noch keine Bewertungen
- Psychology of Face Recognition Brief Introduction 2ndeditionDokument34 SeitenPsychology of Face Recognition Brief Introduction 2ndeditionKevin Brewer100% (6)
- Botanical Oils For Skin CareDokument17 SeitenBotanical Oils For Skin CareTrần NhỏNoch keine Bewertungen
- Soil Classification (Taxonomy)Dokument58 SeitenSoil Classification (Taxonomy)Quinnee VallejosNoch keine Bewertungen
- Myofascial Meridians PDFDokument9 SeitenMyofascial Meridians PDFPedja Ley100% (3)
- "And, When You Want Something, ALL The: UniverseDokument1 Seite"And, When You Want Something, ALL The: UniverseSayNoch keine Bewertungen
- Gases AnswersDokument8 SeitenGases AnswersSayNoch keine Bewertungen
- Chemistry 222 Exam 4: Chapters 11, 13, 14 (16ptsDokument7 SeitenChemistry 222 Exam 4: Chapters 11, 13, 14 (16ptsSayNoch keine Bewertungen
- Element HGDokument3 SeitenElement HGSayNoch keine Bewertungen
- Periodic TableDokument65 SeitenPeriodic Tablerajesh050384Noch keine Bewertungen
- Board of Chemistry-CEDokument2 SeitenBoard of Chemistry-CEElly Paul Andres TomasNoch keine Bewertungen
- Solubility Product ConstantDokument3 SeitenSolubility Product ConstantIka AmbarsariNoch keine Bewertungen
- pKa Chart: Strongest Acids and Weakest BasesDokument2 SeitenpKa Chart: Strongest Acids and Weakest BasesSayNoch keine Bewertungen
- Tables For Group TheoryDokument39 SeitenTables For Group Theoryvitoribeiro90Noch keine Bewertungen
- StemsDokument43 SeitenStemsSayNoch keine Bewertungen
- Spectrum Problem 7Dokument1 SeiteSpectrum Problem 7SayNoch keine Bewertungen
- Spectrum Problem 8Dokument1 SeiteSpectrum Problem 8SayNoch keine Bewertungen
- Spectrum Problem 6Dokument1 SeiteSpectrum Problem 6SayNoch keine Bewertungen
- Spectrum Problem 5Dokument1 SeiteSpectrum Problem 5SayNoch keine Bewertungen
- Spectrum Problem 3Dokument1 SeiteSpectrum Problem 3SayNoch keine Bewertungen
- Spectrum Problem 4Dokument1 SeiteSpectrum Problem 4SayNoch keine Bewertungen
- Spectrum Problem 2Dokument1 SeiteSpectrum Problem 2SayNoch keine Bewertungen
- (Part-I MCQS) (Compulsory)Dokument7 Seiten(Part-I MCQS) (Compulsory)Muzammil RehmanNoch keine Bewertungen
- Birds of Prey PDFDokument24 SeitenBirds of Prey PDFPaula MatosNoch keine Bewertungen
- Lecture # 1 by Amir Afzal Khan: Fundamental ofDokument58 SeitenLecture # 1 by Amir Afzal Khan: Fundamental ofUbaid KhanNoch keine Bewertungen
- Mzbca N2 LK 7Dokument342 SeitenMzbca N2 LK 7Jesús Enrique Turín VilcaNoch keine Bewertungen
- Karakterisasi Koleksi Plasma Nutfah Tomat Lokal Dan IntroduksiDokument7 SeitenKarakterisasi Koleksi Plasma Nutfah Tomat Lokal Dan IntroduksiAlexanderNoch keine Bewertungen
- TATIANADokument8 SeitenTATIANAKaizer NdoloNoch keine Bewertungen
- Body Condition Scoring BCS FinalDokument12 SeitenBody Condition Scoring BCS FinalHamza HarbNoch keine Bewertungen
- Ashton Brennecke CV 2019Dokument3 SeitenAshton Brennecke CV 2019api-456822679Noch keine Bewertungen
- Jhean Steffan Martines de CamargoDokument2 SeitenJhean Steffan Martines de CamargoJheanNoch keine Bewertungen
- Quiz 9 14 Answers PDFDokument2 SeitenQuiz 9 14 Answers PDFюрий локтионовNoch keine Bewertungen
- SWP 331 Final Exam NotesDokument27 SeitenSWP 331 Final Exam Notesdcadenas1Noch keine Bewertungen
- Cell Structure and Function ChapterDokument80 SeitenCell Structure and Function ChapterDrAmit Verma0% (2)
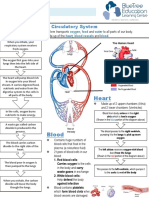
- Circulatory System Notes SLDokument1 SeiteCirculatory System Notes SLRathna PalaniappanNoch keine Bewertungen
- Menthae Piperitae FoliumDokument2 SeitenMenthae Piperitae FoliumartemNoch keine Bewertungen
- 5.2 BiophiliaDokument24 Seiten5.2 BiophiliaNurFarahinZakariahNoch keine Bewertungen
- Pharmacogenetics Drug Drug Interactions1Dokument16 SeitenPharmacogenetics Drug Drug Interactions1Marfu'ah Mar'ahNoch keine Bewertungen
- Individual Differences and Emotional IntelligenceDokument91 SeitenIndividual Differences and Emotional IntelligenceSREEKUTTY K MNoch keine Bewertungen
- Fish Reproduction StrategiesDokument30 SeitenFish Reproduction StrategiesWahyuniFanggiTasikNoch keine Bewertungen
- Campion Mumford Morgeson Nahrgang 2005 PDFDokument24 SeitenCampion Mumford Morgeson Nahrgang 2005 PDFioanaelena89Noch keine Bewertungen
- PDF 4Dokument19 SeitenPDF 4Royyan AdiwijayaNoch keine Bewertungen
- Notes - Growth and DevelopmentDokument4 SeitenNotes - Growth and DevelopmentNathan mwapeNoch keine Bewertungen
- Seroepidemiology of Clamaydia TDokument8 SeitenSeroepidemiology of Clamaydia TMulatuNoch keine Bewertungen
- F94L Is The Limousin Power' GenDokument2 SeitenF94L Is The Limousin Power' GenJose CastanedoNoch keine Bewertungen
- The Digestive System Challenges and OpportunitiesDokument9 SeitenThe Digestive System Challenges and OpportunitiesPhelipe PessanhaNoch keine Bewertungen