Das könnte Ihnen auch gefallen
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Bone Graft Substitutes and Bone Regenerative Engineering 2ed - Cato T. Laurencin, Tao Jiang PDFDokument398 SeitenBone Graft Substitutes and Bone Regenerative Engineering 2ed - Cato T. Laurencin, Tao Jiang PDFJose Andres Noriega100% (1)
- Srotas Final 2Dokument112 SeitenSrotas Final 2Sanjana SajjanarNoch keine Bewertungen
- Anaphy - Chapter-3-Cells-andTissuesDokument80 SeitenAnaphy - Chapter-3-Cells-andTissuesrhmanipol100% (1)
- All About BonesDokument2 SeitenAll About Bonesantonia putriNoch keine Bewertungen
- Ready To Go Lessons For Science Stage 4 Answers PDFDokument9 SeitenReady To Go Lessons For Science Stage 4 Answers PDFSTELLA MARY J100% (2)
- Atlas de Endodoncia 2a Ed. (C. Stock Et Al, Hartcourt Brace 1996) VirgencitadelospxDokument276 SeitenAtlas de Endodoncia 2a Ed. (C. Stock Et Al, Hartcourt Brace 1996) VirgencitadelospxLeidi Diana Marlenne100% (2)
- Anat 106Dokument15 SeitenAnat 106markuspinkusNoch keine Bewertungen
- History of OsteosynthesisDokument9 SeitenHistory of OsteosynthesisPankaj VatsaNoch keine Bewertungen
- Medicinal Properties of Plants From The Genus CissusDokument7 SeitenMedicinal Properties of Plants From The Genus CissusمحمدأميندماجNoch keine Bewertungen
- Aplastic Anemia and HomoeopathyDokument3 SeitenAplastic Anemia and HomoeopathyDr. Rajneesh Kumar Sharma MD Hom100% (1)
- NUR100 Sherpath CH 45 Nutrition ImbalanceDokument4 SeitenNUR100 Sherpath CH 45 Nutrition Imbalancecaloy2345caloyNoch keine Bewertungen
- DLP Science 4 (Human Skeletal System)Dokument2 SeitenDLP Science 4 (Human Skeletal System)Norman NaponeNoch keine Bewertungen
- Implant MaterialsDokument16 SeitenImplant MaterialsNikita AggarwalNoch keine Bewertungen
- Life's Response and Coordination SystemDokument4 SeitenLife's Response and Coordination Systemnik muhiNoch keine Bewertungen
- Link B, Et Al. Orthoped Rev (Pavia) - 2010Dokument9 SeitenLink B, Et Al. Orthoped Rev (Pavia) - 2010Elena CarbonellNoch keine Bewertungen
- Periodontal & Bone Response To Normal Function: 16-4-2020 by DR - Anees MaweriDokument57 SeitenPeriodontal & Bone Response To Normal Function: 16-4-2020 by DR - Anees MaweriAnees MaweriNoch keine Bewertungen
- Chapter-08 - Examination of Musculoskeletal SystemDokument9 SeitenChapter-08 - Examination of Musculoskeletal SystemAnnahi BcNoch keine Bewertungen
- Ncert Page Wise Q Structural OrganizationDokument16 SeitenNcert Page Wise Q Structural OrganizationSagarNoch keine Bewertungen
- FACT Syllabus 15.02.2022Dokument49 SeitenFACT Syllabus 15.02.2022Raju PrinceNoch keine Bewertungen
- AP1 Lab4 Intro To Skeleton FA2021Dokument15 SeitenAP1 Lab4 Intro To Skeleton FA2021Kim VenturaNoch keine Bewertungen
- Progress Test 10A: GrammarDokument6 SeitenProgress Test 10A: Grammarlich dinhNoch keine Bewertungen
- Writing Radiology Reports in ChiropracticDokument6 SeitenWriting Radiology Reports in ChiropracticRohit SagarNoch keine Bewertungen
- Blood Supply of Bones: DR Thouseef A Majeed MS Ortho PG VMKVMCH SalemDokument40 SeitenBlood Supply of Bones: DR Thouseef A Majeed MS Ortho PG VMKVMCH SalemifrazameerNoch keine Bewertungen
- Regeneration and Repair of Periodontal TissuesDokument11 SeitenRegeneration and Repair of Periodontal TissuesAna Maria Montoya GomezNoch keine Bewertungen
- Osteoarthritis: Dr. C. C. VisserDokument85 SeitenOsteoarthritis: Dr. C. C. VisserAhmad Fathira FitraNoch keine Bewertungen
- Technologies in The Treatment of Bone Marrow Edema SyndromeDokument8 SeitenTechnologies in The Treatment of Bone Marrow Edema SyndromeRobert Partarrieu StegmeierNoch keine Bewertungen
- NachamkinDokument425 SeitenNachamkinapi-532187335Noch keine Bewertungen
- What Is Congenital Anomalies?: Anomalies That Occur During Intrauterine Life. Also Called Birth DefectsDokument17 SeitenWhat Is Congenital Anomalies?: Anomalies That Occur During Intrauterine Life. Also Called Birth DefectsSantosh OjhaNoch keine Bewertungen
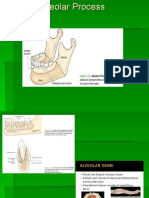
- Alveolar BoneDokument24 SeitenAlveolar BoneHanna SouthwellNoch keine Bewertungen
- Unit 2 SkletonDokument18 SeitenUnit 2 Skletonbertha tandiNoch keine Bewertungen