Das könnte Ihnen auch gefallen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Rosales Vonn Raiven C. M4A PDFDokument3 SeitenRosales Vonn Raiven C. M4A PDFPatrick Jayson VillademosaNoch keine Bewertungen
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
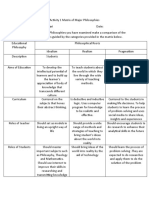
- Activity 1 Matrix of Major PhilosophiesDokument2 SeitenActivity 1 Matrix of Major PhilosophiesHeart Wp100% (1)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Fisip: Jurnal Ilmiah Mahasiswa FISIP Unsyiah Volume 2, Nomor 2: 1-13 Januari 2017 WWW - Jim.unsyiah - ac.id/FISIPDokument13 SeitenFisip: Jurnal Ilmiah Mahasiswa FISIP Unsyiah Volume 2, Nomor 2: 1-13 Januari 2017 WWW - Jim.unsyiah - ac.id/FISIPMusrinNoch keine Bewertungen
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Todes, Body and World PDFDokument385 SeitenTodes, Body and World PDFfirebird4407100% (1)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- MseDokument11 SeitenMseClyde R.OrtegaNoch keine Bewertungen
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- E-RPH 2021: Daily Lesson PlanDokument5 SeitenE-RPH 2021: Daily Lesson Plannel5532Noch keine Bewertungen
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Why Children Talk To Themselves: Laura E. Berk (1994)Dokument7 SeitenWhy Children Talk To Themselves: Laura E. Berk (1994)Ali KalyarNoch keine Bewertungen
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Unit 1 Lesson 1 Assessment 1Dokument2 SeitenUnit 1 Lesson 1 Assessment 1John Rey OñateNoch keine Bewertungen
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- ISTD Training Methodology I August 20 2011Dokument39 SeitenISTD Training Methodology I August 20 2011AnkitBarua100% (1)
- Assessment in Learning 1: Ruel V. Perocho, Maed Sci. Ed. Instructor - BISU Candijay CampusDokument14 SeitenAssessment in Learning 1: Ruel V. Perocho, Maed Sci. Ed. Instructor - BISU Candijay CampusRuel Perocho100% (1)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- For The Full Essay Please WHATSAPP 010-2504287: Assignment / TugasanDokument11 SeitenFor The Full Essay Please WHATSAPP 010-2504287: Assignment / TugasanSimon RajNoch keine Bewertungen
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- Department of Education: Republic of The PhilippinesDokument3 SeitenDepartment of Education: Republic of The PhilippinesJanna GunioNoch keine Bewertungen
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Our Lady of Perpetual Succor CollegeDokument2 SeitenOur Lady of Perpetual Succor CollegeRaven CruizNoch keine Bewertungen
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- Informative SpeechDokument3 SeitenInformative SpeechMaryam KhanzaiNoch keine Bewertungen
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- What Are The 3 Personal Actions or Changes That You Need To Take in Order To Achieve Your Vision of "The Future You"?Dokument2 SeitenWhat Are The 3 Personal Actions or Changes That You Need To Take in Order To Achieve Your Vision of "The Future You"?MAE ANN TOLENTINO100% (1)
- Principles of and Characteristics of Good SpeechDokument3 SeitenPrinciples of and Characteristics of Good SpeechDenz Orbon0% (2)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Principles of Art Design ReportDokument17 SeitenThe Principles of Art Design ReportJane SanNoch keine Bewertungen
- Nur Shahifa BT NorlanDokument2 SeitenNur Shahifa BT NorlanFara Mohd LazimNoch keine Bewertungen
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Learning Jornal 2 - Brenna KingDokument3 SeitenLearning Jornal 2 - Brenna Kingapi-585422168Noch keine Bewertungen
- Grade 1 Addition Lesson PlanDokument3 SeitenGrade 1 Addition Lesson Planapi-539616131Noch keine Bewertungen
- Does The Child Use Visual Function in Most Vision-Related Activities? VFCS Level Identification ChartDokument1 SeiteDoes The Child Use Visual Function in Most Vision-Related Activities? VFCS Level Identification ChartEmiliano Emmanuel RussoNoch keine Bewertungen
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Psych 1XX3 Form Perception II Lecture NotesDokument11 SeitenPsych 1XX3 Form Perception II Lecture NotesGurry MannNoch keine Bewertungen
- Social Learning: by MadhaviDokument12 SeitenSocial Learning: by MadhaviRachamalla KrishnareddyNoch keine Bewertungen
- Albert BanduraDokument5 SeitenAlbert Banduraapi-242298926Noch keine Bewertungen
- 6+1 Trait Beginning Writers GR 1-2Dokument4 Seiten6+1 Trait Beginning Writers GR 1-2lidimessNoch keine Bewertungen
- Activity Sheet: Rosemhier B. Magpantay Bs Psychology 2A (M)Dokument2 SeitenActivity Sheet: Rosemhier B. Magpantay Bs Psychology 2A (M)sandara mallNoch keine Bewertungen
- Attachment Supervisors FormDokument5 SeitenAttachment Supervisors FormawipaT MarowaNoch keine Bewertungen
- Quizz 1 Mkt201Dokument4 SeitenQuizz 1 Mkt201Đạt ĐàoNoch keine Bewertungen
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Daily Lesson Log Personal Dev TDokument34 SeitenDaily Lesson Log Personal Dev TRicky Canico ArotNoch keine Bewertungen
- LP L&S TUTO (Speaking)Dokument5 SeitenLP L&S TUTO (Speaking)TESLA-0619 Jenny Laleng NehNoch keine Bewertungen
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)