Das könnte Ihnen auch gefallen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- Periodic Table PtableDokument1 SeitePeriodic Table Ptableapi-239694539Noch keine Bewertungen
- Partnership LiquidationDokument46 SeitenPartnership LiquidationSewale Abate79% (19)
- Full Download Ebook PDF Introductory Econometrics A Modern Approach 7th Edition by Jeffrey PDFDokument42 SeitenFull Download Ebook PDF Introductory Econometrics A Modern Approach 7th Edition by Jeffrey PDFtimothy.mees27497% (39)
- Activity Based Costing TestbanksDokument18 SeitenActivity Based Costing TestbanksCharlene MinaNoch keine Bewertungen
- Maurice Nicoll The Mark PDFDokument4 SeitenMaurice Nicoll The Mark PDFErwin KroonNoch keine Bewertungen
- Categories of Cargo and Types of ShipsDokument14 SeitenCategories of Cargo and Types of ShipsVibhav Kumar100% (1)
- Learning Module - Joints, Taps and SplicesDokument9 SeitenLearning Module - Joints, Taps and SplicesCarlo Cartagenas100% (1)
- Sofia 2012Dokument2 SeitenSofia 2012MartinMaguNoch keine Bewertungen
- Reaction Mechanism of Coordination ComplexesDokument7 SeitenReaction Mechanism of Coordination ComplexesMartinMagu0% (1)
- Steam Distillation of Essential Oils-Anethole From Anise Followed by Permanganate Oxidation To Anisic AcidDokument1 SeiteSteam Distillation of Essential Oils-Anethole From Anise Followed by Permanganate Oxidation To Anisic AcidMartinMaguNoch keine Bewertungen
- Barometric Hazards Within The Context of Deep-Level Mining: by R.M. Franz and P.C. SchutteDokument4 SeitenBarometric Hazards Within The Context of Deep-Level Mining: by R.M. Franz and P.C. SchutteMartinMaguNoch keine Bewertungen
- V.Santhanam: Department of Chemistry SCSVMVDokument40 SeitenV.Santhanam: Department of Chemistry SCSVMVMartinMaguNoch keine Bewertungen
- Social Studies Notes Class 8 Teacher - Co .KeDokument69 SeitenSocial Studies Notes Class 8 Teacher - Co .KeMartinMaguNoch keine Bewertungen
- Aquasim ManualDokument219 SeitenAquasim Manualpsbc9Noch keine Bewertungen
- Vacancies Advertisement Society Vision: To Redefine Financial WellnessDokument8 SeitenVacancies Advertisement Society Vision: To Redefine Financial WellnessMartinMaguNoch keine Bewertungen
- Steam Distillation of Essential Oils-Anethole From Anise Followed by Permanganate Oxidation To Anisic AcidDokument1 SeiteSteam Distillation of Essential Oils-Anethole From Anise Followed by Permanganate Oxidation To Anisic AcidMartinMaguNoch keine Bewertungen
- Pricelist PDFDokument8 SeitenPricelist PDFMartinMaguNoch keine Bewertungen
- SSHK1PERDokument0 SeitenSSHK1PERitsnotUnUnSUlae24Noch keine Bewertungen
- Mercury in Three Marine Fish SpeciesDokument5 SeitenMercury in Three Marine Fish SpeciesMartinMaguNoch keine Bewertungen
- PostgradOpportunities201516 BrochureDokument38 SeitenPostgradOpportunities201516 BrochureMartinMaguNoch keine Bewertungen
- Under-And Post Graduate Booklet 2014Dokument39 SeitenUnder-And Post Graduate Booklet 2014MartinMaguNoch keine Bewertungen
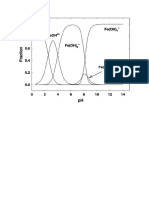
- Fe Speciation With PHDokument2 SeitenFe Speciation With PHMartinMaguNoch keine Bewertungen
- Consular ServicesDokument6 SeitenConsular ServicesMartinMaguNoch keine Bewertungen
- Consular ServicesDokument6 SeitenConsular ServicesMartinMaguNoch keine Bewertungen
- Multimedia University of Kenya: Office of The Estates ManagerDokument2 SeitenMultimedia University of Kenya: Office of The Estates ManagerMartinMaguNoch keine Bewertungen
- Johannes Haushofer's CV of FailuresDokument2 SeitenJohannes Haushofer's CV of FailuresDylan Matthews100% (2)
- Consular ServicesDokument6 SeitenConsular ServicesMartinMaguNoch keine Bewertungen
- 2015-11 List of ERC NRF SA PDFDokument357 Seiten2015-11 List of ERC NRF SA PDFMartinMaguNoch keine Bewertungen
- Determination of Normal Saturated-And Polycyclic Aromatic Hydrocarbons in The River Water of Bangladesh by Liquid-Liquid Extraction and Gas ChromatographyDokument7 SeitenDetermination of Normal Saturated-And Polycyclic Aromatic Hydrocarbons in The River Water of Bangladesh by Liquid-Liquid Extraction and Gas ChromatographyMartinMaguNoch keine Bewertungen
- Ehc 234Dokument256 SeitenEhc 234Daniel BirladeanuNoch keine Bewertungen
- Biochemist 12Dokument5 SeitenBiochemist 12MartinMaguNoch keine Bewertungen
- 1 s2.0 S0301479712001442 MainDokument6 Seiten1 s2.0 S0301479712001442 MainMartinMaguNoch keine Bewertungen
- Multimedia University of Kenya: Office of The Estates ManagerDokument2 SeitenMultimedia University of Kenya: Office of The Estates ManagerMartinMaguNoch keine Bewertungen
- Heavy Metal ToxicityDokument28 SeitenHeavy Metal ToxicityAkpevweoghene Kelvin IdogunNoch keine Bewertungen
- Heat Pyqs NsejsDokument3 SeitenHeat Pyqs NsejsPocketMonTuberNoch keine Bewertungen
- Master Data FileDokument58 SeitenMaster Data Fileinfo.glcom5161Noch keine Bewertungen
- Characteristics of Trochoids and Their Application To Determining Gear Teeth Fillet ShapesDokument14 SeitenCharacteristics of Trochoids and Their Application To Determining Gear Teeth Fillet ShapesJohn FelemegkasNoch keine Bewertungen
- Technical DescriptionDokument2 SeitenTechnical Descriptioncocis_alexandru04995Noch keine Bewertungen
- Newsela Teacher Review - Common Sense EducationDokument1 SeiteNewsela Teacher Review - Common Sense EducationJessicaNoch keine Bewertungen
- State Farm Claims: PO Box 52250 Phoenix AZ 85072-2250Dokument2 SeitenState Farm Claims: PO Box 52250 Phoenix AZ 85072-2250georgia ann polley-yatesNoch keine Bewertungen
- Operator'S Manual PM20X-X-X-BXX: 2" Diaphragm PumpDokument12 SeitenOperator'S Manual PM20X-X-X-BXX: 2" Diaphragm PumpOmar TadeoNoch keine Bewertungen
- Hatayoga 1Dokument11 SeitenHatayoga 1SACHIDANANDA SNoch keine Bewertungen
- H.mohamed Ibrahim Hussain A Study On Technology Updatiing and Its Impact Towards Employee Performance in Orcade Health Care PVT LTD ErodeDokument108 SeitenH.mohamed Ibrahim Hussain A Study On Technology Updatiing and Its Impact Towards Employee Performance in Orcade Health Care PVT LTD ErodeeswariNoch keine Bewertungen
- Model 900 Automated Viscometer: Drilling Fluids EquipmentDokument2 SeitenModel 900 Automated Viscometer: Drilling Fluids EquipmentJazminNoch keine Bewertungen
- RADMASTE CAPS Grade 11 Chemistry Learner GuideDokument66 SeitenRADMASTE CAPS Grade 11 Chemistry Learner Guideamajobe34Noch keine Bewertungen
- R, Axn: Housingand RegulatoryDokument5 SeitenR, Axn: Housingand RegulatoryAce RamosoNoch keine Bewertungen
- NHL DB Rulebook ENGLISHDokument6 SeitenNHL DB Rulebook ENGLISHAdhika WidyaparagaNoch keine Bewertungen
- Wps For Carbon Steel THK 7.11 GtawDokument1 SeiteWps For Carbon Steel THK 7.11 GtawAli MoosaviNoch keine Bewertungen
- Multibody Dynamics Modeling and System Identification For A Quarter-Car Test Rig With McPherson Strut Suspension PDFDokument122 SeitenMultibody Dynamics Modeling and System Identification For A Quarter-Car Test Rig With McPherson Strut Suspension PDFnecromareNoch keine Bewertungen
- Offshore Training Matriz Matriz de Treinamentos OffshoreDokument2 SeitenOffshore Training Matriz Matriz de Treinamentos OffshorecamiladiasmanoelNoch keine Bewertungen
- Rate If Iodine and PropanoneDokument3 SeitenRate If Iodine and Propanoneshareef1Noch keine Bewertungen
- Catch Up RPHDokument6 SeitenCatch Up RPHபிரதீபன் இராதேNoch keine Bewertungen
- Model No. TH-65JX850M/MF Chassis. 9K56T: LED TelevisionDokument53 SeitenModel No. TH-65JX850M/MF Chassis. 9K56T: LED TelevisionRavi ChandranNoch keine Bewertungen
- Alienation Thesis StatementDokument8 SeitenAlienation Thesis Statementafbteepof100% (2)
- 16-ELS-Final-Module 16-08082020Dokument18 Seiten16-ELS-Final-Module 16-08082020jeseca cincoNoch keine Bewertungen
- Salwico CS4000 Fire Detection System: Consilium Marine ABDokument38 SeitenSalwico CS4000 Fire Detection System: Consilium Marine ABJexean SañoNoch keine Bewertungen
- Ce Licensure Examination Problems Rectilinear Translation 6Dokument2 SeitenCe Licensure Examination Problems Rectilinear Translation 6Ginto AquinoNoch keine Bewertungen
- SMMS7 - Mini7 Product Line-Up OverviewDokument46 SeitenSMMS7 - Mini7 Product Line-Up OverviewTín Nguyễn ThànhNoch keine Bewertungen
- Resa Auditing Theorydocx - CompressDokument64 SeitenResa Auditing Theorydocx - CompressMaeNoch keine Bewertungen