Das könnte Ihnen auch gefallen
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (399)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- A510A510M-13 Standard Specification For General Requirements For Wire Rods and Coarse Round Wire, Carbon Steel, and Alloy SteelDokument6 SeitenA510A510M-13 Standard Specification For General Requirements For Wire Rods and Coarse Round Wire, Carbon Steel, and Alloy Steeltjt4779100% (2)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (73)
- Fractional Flow Analysis of WaterfloodDokument11 SeitenFractional Flow Analysis of WaterfloodBagus PrihardianNoch keine Bewertungen
- Physical and Chemical ChangesDokument25 SeitenPhysical and Chemical Changesshreshtha rachakondaNoch keine Bewertungen
- Six Considerations For Proper Dust Collection System DesignDokument9 SeitenSix Considerations For Proper Dust Collection System DesignJoão CoelhoNoch keine Bewertungen
- SKD and SKB Score Comparison TableDokument1 SeiteSKD and SKB Score Comparison TableErikaDwiRahmawatiNoch keine Bewertungen
- Kementerian Agama Republik Indonesia: Jadwal ShalatDokument2 SeitenKementerian Agama Republik Indonesia: Jadwal ShalatErikaDwiRahmawatiNoch keine Bewertungen
- Lampiran P1TLDokument5 SeitenLampiran P1TLErikaDwiRahmawatiNoch keine Bewertungen
- New Microsoft Office PowerPoint PresentationDokument6 SeitenNew Microsoft Office PowerPoint PresentationErikaDwiRahmawatiNoch keine Bewertungen
- Kementerian Agama Republik Indonesia: Jadwal ShalatDokument2 SeitenKementerian Agama Republik Indonesia: Jadwal ShalatErikaDwiRahmawatiNoch keine Bewertungen
- 1 s2.0 S0960894X14007197 MainDokument10 Seiten1 s2.0 S0960894X14007197 MainErikaDwiRahmawatiNoch keine Bewertungen
- Jadwal SholatDokument2 SeitenJadwal SholatErikaDwiRahmawatiNoch keine Bewertungen
- Tractus Digestivus: DR Aris Prasetyo, MkesDokument51 SeitenTractus Digestivus: DR Aris Prasetyo, MkesErikaDwiRahmawatiNoch keine Bewertungen
- Pain Physiology 2015Dokument34 SeitenPain Physiology 2015ErikaDwiRahmawatiNoch keine Bewertungen
- Tractus Digestivus: DR Aris Prasetyo, MkesDokument51 SeitenTractus Digestivus: DR Aris Prasetyo, MkesErikaDwiRahmawatiNoch keine Bewertungen
- AOAC 965.08 Kadar Air Pada PupukDokument3 SeitenAOAC 965.08 Kadar Air Pada PupukDENI ARIFIYANTONoch keine Bewertungen
- Advantages and Disadvantages of Stainless Steel CookwareDokument15 SeitenAdvantages and Disadvantages of Stainless Steel CookwareFaIz FauziNoch keine Bewertungen
- Cationic DyesDokument3 SeitenCationic Dyesmayree_gNoch keine Bewertungen
- FastLUBE lubricants guide for anti-galling and corrosion protectionDokument1 SeiteFastLUBE lubricants guide for anti-galling and corrosion protectionneubofren600Noch keine Bewertungen
- 221 Chloride and SulfateDokument1 Seite221 Chloride and SulfatesofianesedkaouiNoch keine Bewertungen
- Book Print Gunastar Final - 1528261270Dokument103 SeitenBook Print Gunastar Final - 1528261270santoshNoch keine Bewertungen
- Algebra:: N N N N N N 1B 2B ZoneDokument6 SeitenAlgebra:: N N N N N N 1B 2B Zonevenedick sapongayNoch keine Bewertungen
- An Introduction To Vibration Control in BuildingsDokument45 SeitenAn Introduction To Vibration Control in Buildingsosama alabsiNoch keine Bewertungen
- Cutting Fluids and Machining ProcessesDokument19 SeitenCutting Fluids and Machining Processescasey rimNoch keine Bewertungen
- HLB Values and Saa Functionality: Technical Brief 2010 Volume 1Dokument2 SeitenHLB Values and Saa Functionality: Technical Brief 2010 Volume 1ArangaNoch keine Bewertungen
- Aisi 321 Stainless Steel Uns s32100 Din 1.4541Dokument3 SeitenAisi 321 Stainless Steel Uns s32100 Din 1.4541Shariq KhanNoch keine Bewertungen
- Physical Science 20 - Lesson PlanDokument7 SeitenPhysical Science 20 - Lesson Planapi-349567441Noch keine Bewertungen
- Preparing, Cleaning, and Evaluating Corrosion Test SpecimensDokument8 SeitenPreparing, Cleaning, and Evaluating Corrosion Test SpecimensLê CôngNoch keine Bewertungen
- Assignment 1Dokument4 SeitenAssignment 1Nur AfiqahNoch keine Bewertungen
- TDS - RheoFIT 764Dokument2 SeitenTDS - RheoFIT 764aahtagoNoch keine Bewertungen
- Titration Analysis of VinegarDokument2 SeitenTitration Analysis of VinegarSourabh DasNoch keine Bewertungen
- Macromolecules: Question Paper 1Dokument8 SeitenMacromolecules: Question Paper 1Bridgette LombardNoch keine Bewertungen
- PQCDokument77 SeitenPQCAnkit MittalNoch keine Bewertungen
- Determination of Vinyl Chloride Monomer in Food Contact Materials by Solid Phase Microextraction Coupled With Gas Chromatography/Mass SpectrometryDokument6 SeitenDetermination of Vinyl Chloride Monomer in Food Contact Materials by Solid Phase Microextraction Coupled With Gas Chromatography/Mass SpectrometryPhuongthuy127Noch keine Bewertungen
- SP 360 CmanualDokument35 SeitenSP 360 CmanualencontreelrepuestoNoch keine Bewertungen
- Inorganic 3Dokument7 SeitenInorganic 3Ephraim PeasahNoch keine Bewertungen
- RSBV Tech BulletinpdfDokument16 SeitenRSBV Tech BulletinpdfbalancedrumNoch keine Bewertungen
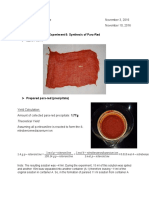
- Synthesis of Para-Red and 1-Iodo-4-Nitrobenzene ExperimentDokument2 SeitenSynthesis of Para-Red and 1-Iodo-4-Nitrobenzene ExperimentMaryNoch keine Bewertungen
- C5Dokument4 SeitenC5Vilma Puma InostrozaNoch keine Bewertungen
- Inlab MC X5 Accessories: Order FormDokument1 SeiteInlab MC X5 Accessories: Order FormCiprian SarivanNoch keine Bewertungen
- The Reactivity Series NotesDokument8 SeitenThe Reactivity Series NotesTHEVINESHNoch keine Bewertungen