Das könnte Ihnen auch gefallen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Gas Chromatography Mass SpectrometryDokument20 SeitenGas Chromatography Mass SpectrometryNirmal SharmaNoch keine Bewertungen
- Encyclopedia of Hormones PDFDokument2.120 SeitenEncyclopedia of Hormones PDFWisnu Adryanto0% (2)
- Lab Report Animal and Plant CellsDokument5 SeitenLab Report Animal and Plant CellsNicolas Gómez100% (1)
- Total Imprest Fund Has Received From GM FDokument1 SeiteTotal Imprest Fund Has Received From GM FNirmal SharmaNoch keine Bewertungen
- SK-078 OSP For ProcessingDokument9 SeitenSK-078 OSP For ProcessingNirmal SharmaNoch keine Bewertungen
- Supermarket Requirement 1Dokument2 SeitenSupermarket Requirement 1Nirmal SharmaNoch keine Bewertungen
- High Performance Liquid Chromatography - HPLCDokument9 SeitenHigh Performance Liquid Chromatography - HPLCNirmal SharmaNoch keine Bewertungen
- Ancient History MCQ PDF by Study Like A ProDokument44 SeitenAncient History MCQ PDF by Study Like A ProNirmal SharmaNoch keine Bewertungen
- Horticulture ICAR JRF Previous Yeras QuestionsDokument12 SeitenHorticulture ICAR JRF Previous Yeras QuestionsNirmal SharmaNoch keine Bewertungen
- Medicine & VedasDokument11 SeitenMedicine & VedasNirmal SharmaNoch keine Bewertungen
- Pursuit of Innovation in VedicDokument18 SeitenPursuit of Innovation in VedicNirmal SharmaNoch keine Bewertungen
- Polymeric MaterialDokument1 SeitePolymeric MaterialNirmal SharmaNoch keine Bewertungen
- June 2018: Monthly Current AffairsDokument74 SeitenJune 2018: Monthly Current AffairsNirmal SharmaNoch keine Bewertungen
- Food Technology & Processing: Introduction To The Microbiology of FoodDokument22 SeitenFood Technology & Processing: Introduction To The Microbiology of FoodNirmal SharmaNoch keine Bewertungen
- Food Preservation by FermentationDokument6 SeitenFood Preservation by FermentationNirmal SharmaNoch keine Bewertungen
- Reproductive System & Sexual Disorders: Current Research: Asexuality: Dysfunction or Sexual Orientation?Dokument3 SeitenReproductive System & Sexual Disorders: Current Research: Asexuality: Dysfunction or Sexual Orientation?Nirmal SharmaNoch keine Bewertungen
- Prohibitory Order 4-4-2020Dokument2 SeitenProhibitory Order 4-4-2020Nirmal SharmaNoch keine Bewertungen
- Thank You For Your Purchase!: Chhapaak (U)Dokument2 SeitenThank You For Your Purchase!: Chhapaak (U)Nirmal SharmaNoch keine Bewertungen
- Milk and Milk ProductsDokument10 SeitenMilk and Milk ProductsNirmal SharmaNoch keine Bewertungen
- DNA and RNA Comparison WorksheetDokument4 SeitenDNA and RNA Comparison WorksheetH DNoch keine Bewertungen
- 6-Translocation in The PhloemDokument35 Seiten6-Translocation in The PhloemSultanah Khaidoo-AubdoollahNoch keine Bewertungen
- Hyprt Doc 214Dokument3 SeitenHyprt Doc 214api-490638499Noch keine Bewertungen
- 2012 EMA Guideline On The Investigation of Drug InteractionsDokument60 Seiten2012 EMA Guideline On The Investigation of Drug InteractionsAnna Margarita BernardoNoch keine Bewertungen
- (Prof. Dr. Klaus Wolf (Auth.) ) Nonconventional Yea (B-Ok - Xyz)Dokument630 Seiten(Prof. Dr. Klaus Wolf (Auth.) ) Nonconventional Yea (B-Ok - Xyz)Muhammad HusseinNoch keine Bewertungen
- Dwnload Full Animal Physiology From Genes To Organisms 2nd Edition Sherwood Test Bank PDFDokument36 SeitenDwnload Full Animal Physiology From Genes To Organisms 2nd Edition Sherwood Test Bank PDFsiphilisdysluite7xrxc100% (14)
- Glycogen Storage Capacity and de Novo Iipogenesis During Massive Carbohydrate Overfeeding in ManDokument9 SeitenGlycogen Storage Capacity and de Novo Iipogenesis During Massive Carbohydrate Overfeeding in ManSalvio EspositoNoch keine Bewertungen
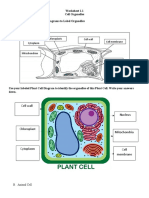
- Worksheet 1.1 Cell OrganelleDokument2 SeitenWorksheet 1.1 Cell OrganelleCyndel TindoyNoch keine Bewertungen
- Medical Progress: Review ArticlesDokument15 SeitenMedical Progress: Review ArticlesFeri EspinosaSerranoNoch keine Bewertungen
- Genetic RecombinationDokument55 SeitenGenetic RecombinationRiyaniMartyasariNoch keine Bewertungen
- Cell Organelles DefinitionDokument40 SeitenCell Organelles Definitionmuhammad aslam0% (1)
- Bigdye Terminator V3.1 Cycle Sequencing Kit: ProtocolDokument72 SeitenBigdye Terminator V3.1 Cycle Sequencing Kit: Protocoliis faridahNoch keine Bewertungen
- Diabetes Mellitus MCQsDokument4 SeitenDiabetes Mellitus MCQsvaidehianand007Noch keine Bewertungen
- Overview of Post-Translational Modification AnalysisDokument2 SeitenOverview of Post-Translational Modification AnalysisSteven4654Noch keine Bewertungen
- Nucleic Acids HandoutDokument3 SeitenNucleic Acids HandoutAlifah SyarafinaNoch keine Bewertungen
- BioinformaticsDokument10 SeitenBioinformaticsRamla FatimaNoch keine Bewertungen
- Dietrich 2017Dokument10 SeitenDietrich 2017Zulfi Nur Amrina RosyadaNoch keine Bewertungen
- AB BC : Part ADokument39 SeitenAB BC : Part AApoorva GuptaNoch keine Bewertungen
- Biomedical Importance Glycolysis Can Function Under Anaerobic ConditionsDokument5 SeitenBiomedical Importance Glycolysis Can Function Under Anaerobic ConditionsCatalina Denise Blaquera FloresNoch keine Bewertungen
- BS1001 LipidsDokument32 SeitenBS1001 LipidsLiloyi LupiyaNoch keine Bewertungen
- POLYSACCHARIDESDokument2 SeitenPOLYSACCHARIDESYholzManioNoch keine Bewertungen
- (Pds Patklin) : Perhimpunan Dokter Spesialis Patologi Klinik Dan Kedokteran Laboratorium IndonesiaDokument13 Seiten(Pds Patklin) : Perhimpunan Dokter Spesialis Patologi Klinik Dan Kedokteran Laboratorium IndonesiaNida ChoerunnisaNoch keine Bewertungen
- Ligand ProteinDokument4 SeitenLigand ProteinOscar George LuNoch keine Bewertungen
- IB Biology SL Topics Study GuideDokument65 SeitenIB Biology SL Topics Study GuideSai 0235Noch keine Bewertungen
- Menselijke Biologie en Ziekteleer SamenvattingDokument167 SeitenMenselijke Biologie en Ziekteleer SamenvattingLotte WoutersNoch keine Bewertungen
- Mechanism of Gastric SecretionDokument11 SeitenMechanism of Gastric Secretionmulya ningsihNoch keine Bewertungen
- Test Series For Neet-2020Dokument11 SeitenTest Series For Neet-2020kavyareddyNoch keine Bewertungen
- Pure Bio CH 5 Textbook Answers PDFDokument2 SeitenPure Bio CH 5 Textbook Answers PDFMahamIsmailNoch keine Bewertungen