Das könnte Ihnen auch gefallen
- 06 - Retículo EndoplasmáticoDokument55 Seiten06 - Retículo EndoplasmáticocesarjbcNoch keine Bewertungen
- Semana 3 - BASES MOLECULARES 3Dokument25 SeitenSemana 3 - BASES MOLECULARES 3Alison YalleNoch keine Bewertungen
- Fundamentos de Fisiología - Ensayo 2Dokument9 SeitenFundamentos de Fisiología - Ensayo 2Roberto JimenezNoch keine Bewertungen
- CemorDokument8 SeitenCemorclaudioNoch keine Bewertungen
- Reticulo EndoplasmaticoDokument9 SeitenReticulo EndoplasmaticoEnrique Ochoa0% (1)
- 8.sistemas Membranosos de La Célula EucariotaDokument9 Seiten8.sistemas Membranosos de La Célula EucariotaAntony VillamarNoch keine Bewertungen
- La Celula, ImprimirDokument39 SeitenLa Celula, ImprimirEstay A DanielNoch keine Bewertungen
- Informe EditadoDokument27 SeitenInforme EditadoErick SaldañaNoch keine Bewertungen
- Sistema de EndomembranasDokument9 SeitenSistema de Endomembranasandrea rochaNoch keine Bewertungen
- Sistema de EndomembranasDokument16 SeitenSistema de EndomembranasAracelyIssaEnriquezTorrezNoch keine Bewertungen
- Exposicion 12Dokument11 SeitenExposicion 12CAMILA JELENA ESTRADA ALVARADONoch keine Bewertungen
- Expo SISTEMA MEMBRANASDokument28 SeitenExpo SISTEMA MEMBRANASVANIA ALEXA COTRINA ESTEVESNoch keine Bewertungen
- CELULADokument108 SeitenCELULAalejandraNoch keine Bewertungen
- 1 - La Célula y Sus Funciones - URLDokument45 Seiten1 - La Célula y Sus Funciones - URLPablo GonzalezNoch keine Bewertungen
- Informe N 5 BiologiaDokument9 SeitenInforme N 5 BiologiaskinlowNoch keine Bewertungen
- Organelos CelularesDokument9 SeitenOrganelos CelularesLuz Ang�lica Sandoval S�nchezNoch keine Bewertungen
- Componentes de La CelulaDokument5 SeitenComponentes de La CelulaMarco Antonio Quiroga BustinzaNoch keine Bewertungen
- Trabajo de BiologiaDokument11 SeitenTrabajo de Biologialoaizaj12Noch keine Bewertungen
- Capitulo 12 AlbertsDokument35 SeitenCapitulo 12 AlbertsRicardo Cuatlayotl OlarteNoch keine Bewertungen
- Eucariota, Organelas, Citoplasma, CitoesqueletoDokument30 SeitenEucariota, Organelas, Citoplasma, CitoesqueletoAlan GanemNoch keine Bewertungen
- Resumen OATec - Biologia MolecularDokument32 SeitenResumen OATec - Biologia MolecularLucia LlordenNoch keine Bewertungen
- Celula, Morfologia CelularDokument8 SeitenCelula, Morfologia Celularscarletlugo4751Noch keine Bewertungen
- Tráfico VesicularDokument55 SeitenTráfico VesicularAficionadoNoch keine Bewertungen
- Resumen Biología CBCDokument35 SeitenResumen Biología CBCNumilén PiqueroNoch keine Bewertungen
- BIOLOGÍA Tema 9 (Bloque 2)Dokument7 SeitenBIOLOGÍA Tema 9 (Bloque 2)Carolina De la peña fernandezNoch keine Bewertungen
- ReticuloDokument9 SeitenReticuloAngeles Jacqueline Esparza CerinoNoch keine Bewertungen
- Organitos CitoplasmaticosDokument4 SeitenOrganitos CitoplasmaticosMartNoch keine Bewertungen
- Citoplasma y NucleoDokument32 SeitenCitoplasma y NucleoAngela C. ChavezNoch keine Bewertungen
- Copia de Sistemas de EndomembranasDokument8 SeitenCopia de Sistemas de EndomembranasMartina AgueroNoch keine Bewertungen
- Organelos CelularesDokument38 SeitenOrganelos Celularesyoselin gonzalezNoch keine Bewertungen
- Practica de Biologia Semana 11Dokument9 SeitenPractica de Biologia Semana 11Fernando LiviasNoch keine Bewertungen
- Guia de Apoyo BioquimicaDokument8 SeitenGuia de Apoyo BioquimicaCarla Cecilia Gonzalez SuberoNoch keine Bewertungen
- Resumen Del Capitulo 2 y 3 Guyton y HallDokument7 SeitenResumen Del Capitulo 2 y 3 Guyton y HallEdward ArredondoNoch keine Bewertungen
- Reticulo Endoplasmatico. Grupo 6Dokument9 SeitenReticulo Endoplasmatico. Grupo 6XxTAXIXLOLxXNoch keine Bewertungen
- 3-La CelulaDokument5 Seiten3-La CelulaVICTOR FUENTESNoch keine Bewertungen
- Modulo 8-04Dokument17 SeitenModulo 8-04Gustavo RaeleNoch keine Bewertungen
- La Celula y Elementos ConstitutivosDokument45 SeitenLa Celula y Elementos ConstitutivosBeth DiazNoch keine Bewertungen
- EnsayoDokument7 SeitenEnsayoDiana Vite RomeroNoch keine Bewertungen
- Retículo EndoplasmáticoDokument8 SeitenRetículo Endoplasmáticocongaconga123Noch keine Bewertungen
- Fisiologia FinalDokument7 SeitenFisiologia FinalDaniela VelascoNoch keine Bewertungen
- Clase 7 - CélulasDokument16 SeitenClase 7 - CélulasDafne CarballoNoch keine Bewertungen
- Diversidad CelularDokument40 SeitenDiversidad CelularNiebla001Noch keine Bewertungen
- Reticulo Endoplasmatico Liso y Rugoso, FuncionesDokument5 SeitenReticulo Endoplasmatico Liso y Rugoso, FuncionesAna MarcelaNoch keine Bewertungen
- Tema10 Orgánulos Membranosos..Dokument18 SeitenTema10 Orgánulos Membranosos..Duveily AcostaNoch keine Bewertungen
- Organelos CitoplasmáticosDokument6 SeitenOrganelos CitoplasmáticosMaribel Zavala100% (1)
- Histologia OrganoidesDokument51 SeitenHistologia OrganoidesDaniel Taya AvalosNoch keine Bewertungen
- Celula Eucariota. OrganelasDokument37 SeitenCelula Eucariota. Organelasjesus espejoNoch keine Bewertungen
- EucariotasDokument49 SeitenEucariotasMohamed AléNoch keine Bewertungen
- Unidad IV. MembranaDokument57 SeitenUnidad IV. Membranamariangeles292003Noch keine Bewertungen
- 24 EndomembranaDokument4 Seiten24 EndomembranaFreddy David HerreraNoch keine Bewertungen
- Celula Animal02Dokument43 SeitenCelula Animal02NAYELLY DE LOS ANGELES GONZA GUEVARANoch keine Bewertungen
- La Celula AnimalDokument9 SeitenLa Celula AnimalJuly CollantesNoch keine Bewertungen
- Marco TeóricoDokument6 SeitenMarco TeóricoEdison OrtizNoch keine Bewertungen
- NUCLEODokument10 SeitenNUCLEOCarolina MachucaNoch keine Bewertungen
- La Estructura de La CélulaDokument6 SeitenLa Estructura de La CélulaMaricielo Calderon MinayaNoch keine Bewertungen
- Unida-2 Semana 9 Clase 2 MiercolesDokument68 SeitenUnida-2 Semana 9 Clase 2 Miercolesbriyith garciaNoch keine Bewertungen
- Consultas de Biologia Estructura de La Celula VegetalDokument15 SeitenConsultas de Biologia Estructura de La Celula VegetalMARIBEL DELGADONoch keine Bewertungen
- Sistema de EndomembranasDokument35 SeitenSistema de EndomembranasMaria Paula Quintero TamayoNoch keine Bewertungen
- Informe Reticulo EndoplasmaticoDokument9 SeitenInforme Reticulo EndoplasmaticoJosse RodriguezNoch keine Bewertungen
- QUE ES El Cierre de VentasDokument2 SeitenQUE ES El Cierre de VentasGil RuanoNoch keine Bewertungen
- El Cierre de La VentaDokument4 SeitenEl Cierre de La VentaGil RuanoNoch keine Bewertungen
- Cómo Hacer Un Cierre de Ventas ExitosoDokument7 SeitenCómo Hacer Un Cierre de Ventas ExitosoGil RuanoNoch keine Bewertungen
- Las 7 Mejores Técnicas de Cierre de VentasDokument7 SeitenLas 7 Mejores Técnicas de Cierre de VentasGil RuanoNoch keine Bewertungen
- 13 Técnicas de Cierre de VentasDokument8 Seiten13 Técnicas de Cierre de VentasGil RuanoNoch keine Bewertungen
- Cierre de VentasDokument4 SeitenCierre de VentasGil RuanoNoch keine Bewertungen
- Tipos de CÃ - Lulas MadreDokument9 SeitenTipos de CÃ - Lulas MadreGil RuanoNoch keine Bewertungen
- Tecnicas Estrategias Pensamiento Critico Maureen Priestley PDFDokument41 SeitenTecnicas Estrategias Pensamiento Critico Maureen Priestley PDFGil RuanoNoch keine Bewertungen
- Prob Simulacion2005 2006Dokument6 SeitenProb Simulacion2005 2006Gil RuanoNoch keine Bewertungen
- Bioqumica2 140307082934 Phpapp01Dokument21 SeitenBioqumica2 140307082934 Phpapp01Julimar FloresNoch keine Bewertungen
- Ejercicios Hiragana PDFDokument10 SeitenEjercicios Hiragana PDFGJe001Noch keine Bewertungen
- Agua, Electrolitos y Equilibrio Ácido-Base - Ayus PDFDokument512 SeitenAgua, Electrolitos y Equilibrio Ácido-Base - Ayus PDFPam Moyolema100% (4)
- Manual Del MaestroDokument64 SeitenManual Del MaestroNozomi Fudo Ishi90% (29)
- Normas ApaDokument32 SeitenNormas ApaGil RuanoNoch keine Bewertungen
- Medidas de Prevencion Y Control Ante Un BroteDokument20 SeitenMedidas de Prevencion Y Control Ante Un BroteGil RuanoNoch keine Bewertungen
- Sistema UrinarioDokument85 SeitenSistema UrinarioGil RuanoNoch keine Bewertungen
- Trabajo de AmnesiaDokument8 SeitenTrabajo de AmnesiaGil RuanoNoch keine Bewertungen
- Investigacion de Brotes Leccion6Dokument60 SeitenInvestigacion de Brotes Leccion6Gil RuanoNoch keine Bewertungen
- Normas ApaDokument32 SeitenNormas ApaGil RuanoNoch keine Bewertungen
- Curso Química 2015 4 Tabla y Ley PeriódicaDokument131 SeitenCurso Química 2015 4 Tabla y Ley PeriódicaGil RuanoNoch keine Bewertungen
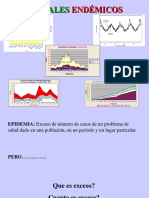
- Corredor Endemico Clase 10 AbrilDokument24 SeitenCorredor Endemico Clase 10 AbrilGil RuanoNoch keine Bewertungen
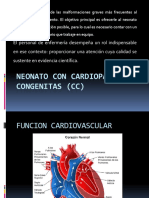
- Neonato Con Cardiopatías Congenitas (CC)Dokument42 SeitenNeonato Con Cardiopatías Congenitas (CC)Gil RuanoNoch keine Bewertungen
- 300 LibrosparaleerDokument128 Seiten300 LibrosparaleerAle Bibliotecaria0% (1)
- Historia Del FútbolDokument3 SeitenHistoria Del FútbolGil RuanoNoch keine Bewertungen
- Medicion 1Dokument29 SeitenMedicion 1Gil RuanoNoch keine Bewertungen
- 03 Still Just HumanDokument90 Seiten03 Still Just HumanGil RuanoNoch keine Bewertungen
- Ejercicio de IndicadoresDokument1 SeiteEjercicio de IndicadoresGil RuanoNoch keine Bewertungen
- Ejercicio de IndicadoresDokument1 SeiteEjercicio de IndicadoresGil RuanoNoch keine Bewertungen
- 02still HumanDokument253 Seiten02still HumanGil Ruano100% (1)
- 01just HumanDokument254 Seiten01just HumanGil RuanoNoch keine Bewertungen
- Manual Bio Cel. 2018Dokument132 SeitenManual Bio Cel. 2018Juanes Gallego0% (1)
- Guía Biología Sexto 2° Periodo Marcela ZapataDokument10 SeitenGuía Biología Sexto 2° Periodo Marcela ZapataAlexa SotoNoch keine Bewertungen
- Biología CelularDokument93 SeitenBiología CelularAlejandra GonzalezNoch keine Bewertungen
- Preguntas Parcial 1. Biologia Celular y Molecular.Dokument4 SeitenPreguntas Parcial 1. Biologia Celular y Molecular.MARIANA ALDANA CASTELLANOSNoch keine Bewertungen
- 23 CelulasEucariotasDokument71 Seiten23 CelulasEucariotasaureliano_fernándezNoch keine Bewertungen
- Fase 1 - Conocer La Estructura de La Célula Vegetal, 1 Darwin CastilloDokument13 SeitenFase 1 - Conocer La Estructura de La Célula Vegetal, 1 Darwin Castillodarwin alfonso castillo100% (1)
- Partes de La CélulaDokument16 SeitenPartes de La CélulaChristian RodriguezNoch keine Bewertungen
- Organelos CelularesDokument20 SeitenOrganelos CelularesEspinoza Flores Jesús ANoch keine Bewertungen
- Las Organelas Celulares y Su FunciónDokument14 SeitenLas Organelas Celulares y Su FunciónMaria LoraNoch keine Bewertungen
- Biologia JHDokument5 SeitenBiologia JHLuis Levano HernandezNoch keine Bewertungen
- Sistemas Endomembranas, Citoesqueleto.Dokument38 SeitenSistemas Endomembranas, Citoesqueleto.andreafonnegraNoch keine Bewertungen
- CITOESTRUCTURA Histologia BiologiaDokument47 SeitenCITOESTRUCTURA Histologia BiologiaSilvia SiacaraNoch keine Bewertungen
- Biologia Selectividad Examen 11 Resuelto Castilla La Mancha WWW - Siglo21x.blogspotDokument9 SeitenBiologia Selectividad Examen 11 Resuelto Castilla La Mancha WWW - Siglo21x.blogspotmonografiasx100% (1)
- (ppt6) Compartimientos y Transporte Intracelular 1 y 2 PDFDokument8 Seiten(ppt6) Compartimientos y Transporte Intracelular 1 y 2 PDFcamila palomino100% (2)
- Apunte Vacuolas y Sustancias ErgásticasDokument9 SeitenApunte Vacuolas y Sustancias ErgásticasJovana Jai'meNoch keine Bewertungen
- GUIA 5 BIO 035 UnabDokument13 SeitenGUIA 5 BIO 035 UnabJoséTomasSwettBarrosNoch keine Bewertungen
- Biología - Tarea 2 - Grupo 30176 - 64Dokument25 SeitenBiología - Tarea 2 - Grupo 30176 - 64Ana Lucia CardonaNoch keine Bewertungen
- Célula EucariotaDokument55 SeitenCélula EucariotaErnesto Lluesma DropoNoch keine Bewertungen
- Membrana CitoplasmaticaDokument13 SeitenMembrana CitoplasmaticaLuisaFernanda Aroca OlivellaNoch keine Bewertungen
- Kinesiologia BiologiaDokument12 SeitenKinesiologia BiologiaExequiel Pereyra100% (1)
- Control N°2 Biología Mención Preu7 PDFDokument9 SeitenControl N°2 Biología Mención Preu7 PDFMidari ChinchillaNoch keine Bewertungen
- Citoplasma Celular FinalDokument8 SeitenCitoplasma Celular FinaljulipeceNoch keine Bewertungen
- GEOBIOLOGIA PrevencionDokument87 SeitenGEOBIOLOGIA Prevencionminunbox100% (6)
- CELULA 2-CepreDokument78 SeitenCELULA 2-CepreHarumi Montalvan VegaNoch keine Bewertungen
- Semana 5 - Membranas Biologicas, Citoplasma y CitoesqueletoDokument95 SeitenSemana 5 - Membranas Biologicas, Citoplasma y CitoesqueletoLuis MEZA MACHADONoch keine Bewertungen
- Biologia Celular y Genetica 2017Dokument10 SeitenBiologia Celular y Genetica 2017sebastian floresNoch keine Bewertungen
- Histología: Atlas en Color y Texto deDokument33 SeitenHistología: Atlas en Color y Texto decarolinaNoch keine Bewertungen
- 2.1.-Composición de Celular 1Dokument6 Seiten2.1.-Composición de Celular 1María Espinosa de los MonterosNoch keine Bewertungen
- Resumen BiologiaDokument90 SeitenResumen BiologiaDafne Olenka Davila PerezNoch keine Bewertungen
- Tesis Doctoral MC Chiale - compressed.pdf-PDFA PDFDokument229 SeitenTesis Doctoral MC Chiale - compressed.pdf-PDFA PDFElizaabethNoch keine Bewertungen