Das könnte Ihnen auch gefallen
- Tema 3Dokument4 SeitenTema 3agzestvzNoch keine Bewertungen
- Los Lípidos OfficialDokument20 SeitenLos Lípidos OfficialLondoño J AlejandroNoch keine Bewertungen
- Lipidos 2Dokument6 SeitenLipidos 2mattyurrutia17Noch keine Bewertungen
- BIOQUIMICADokument5 SeitenBIOQUIMICAMaria Jose Cañizarez JimenezNoch keine Bewertungen
- Tarea LipidosDokument3 SeitenTarea LipidosYessi HdezNoch keine Bewertungen
- TEMA 5 LípidosDokument5 SeitenTEMA 5 LípidosElena Sabas CarvajalNoch keine Bewertungen
- Tarea LipidosDokument3 SeitenTarea LipidosYessi HdezNoch keine Bewertungen
- LipidosDokument14 SeitenLipidosCecilia Márquez MoralesNoch keine Bewertungen
- 03 LípidosDokument7 Seiten03 LípidosbeabergamoNoch keine Bewertungen
- LÍPIDOSDokument4 SeitenLÍPIDOSwert addNoch keine Bewertungen
- LipidosDokument13 SeitenLipidosSarita Herrera MartinezNoch keine Bewertungen
- Química de LípidosDokument38 SeitenQuímica de LípidosJIMMY HUMBERTO IZA IZANoch keine Bewertungen
- Lipidos Trabajo PracticoDokument20 SeitenLipidos Trabajo PracticoCristhian Jose Tejada Merino100% (1)
- Lipidos ExpoDokument9 SeitenLipidos Expoedu_manuel1995Noch keine Bewertungen
- LIPIDOSDokument4 SeitenLIPIDOSCesar DavalosNoch keine Bewertungen
- Lípidos: definición, clasificación, ácidos grasos y síntesisDokument8 SeitenLípidos: definición, clasificación, ácidos grasos y síntesisPier MndzNoch keine Bewertungen
- Erick Jean Pierre Mendoza VasquezDokument10 SeitenErick Jean Pierre Mendoza VasquezPier MndzNoch keine Bewertungen
- LIPIDOSDokument10 SeitenLIPIDOSlourdessbravooNoch keine Bewertungen
- Reporte LipidosDokument20 SeitenReporte Lipidosjunieth sequeiraNoch keine Bewertungen
- Lípidos 20 - 06Dokument34 SeitenLípidos 20 - 06Alfredo MéndezNoch keine Bewertungen
- Diapositivas de LipidosDokument54 SeitenDiapositivas de LipidosELKIN ARMEJO GARCIA MENDEZNoch keine Bewertungen
- Fosfolipidos y GlucolipidosDokument8 SeitenFosfolipidos y Glucolipidosrafael ramirezNoch keine Bewertungen
- 4 LipidosDokument15 Seiten4 LipidosAndres Felipe Prieto AlarconNoch keine Bewertungen
- Aspectos Generales de Los LípidosDokument14 SeitenAspectos Generales de Los LípidosmarcelaNoch keine Bewertungen
- Lípidos: concepto, funciones y clasificaciónDokument7 SeitenLípidos: concepto, funciones y clasificaciónPaula Sánchez JiménezNoch keine Bewertungen
- Aspectos Generales de Los LípidosDokument4 SeitenAspectos Generales de Los LípidospanteraggNoch keine Bewertungen
- Copia de 7. LÍPIDOS - EdumedDokument17 SeitenCopia de 7. LÍPIDOS - EdumedCamz NagoNoch keine Bewertungen
- Benemerita Universidad Autonoma de PueblaDokument7 SeitenBenemerita Universidad Autonoma de PueblaDiagsa MktNoch keine Bewertungen
- Informe de Práctica de LipidosDokument4 SeitenInforme de Práctica de LipidosmaalhuesNoch keine Bewertungen
- T 4. - LípidosDokument61 SeitenT 4. - Lípidosandreacastillo22154Noch keine Bewertungen
- Lípidos: Características, clasificación y funcionesDokument3 SeitenLípidos: Características, clasificación y funcionesAnaNoch keine Bewertungen
- Guia de Estudio - MacromoleculasDokument22 SeitenGuia de Estudio - MacromoleculasFlor GonzalezNoch keine Bewertungen
- Practica No. 5. LipidosDokument9 SeitenPractica No. 5. LipidosDaniel Villabona BeltranNoch keine Bewertungen
- Triptico de Lípidos 1Dokument2 SeitenTriptico de Lípidos 1Ymer Altamirano Diaz78% (9)
- Lípidos: funciones y clasificaciónDokument46 SeitenLípidos: funciones y clasificaciónJhanneth Choque MuruchiNoch keine Bewertungen
- Taller de Lipidos ...Dokument4 SeitenTaller de Lipidos ...Jhonatan IbarraNoch keine Bewertungen
- LIPIDOSDokument5 SeitenLIPIDOSfrancisca menaNoch keine Bewertungen
- 2ºbachillerato, Tema 3 (Lípidos)Dokument7 Seiten2ºbachillerato, Tema 3 (Lípidos)ms marvelNoch keine Bewertungen
- LÍPIDOSDokument3 SeitenLÍPIDOSVicentino LópezNoch keine Bewertungen
- Lipidos y Acidos Nucleicos (Trabajo) - Del Aguila Soto, OgerDokument87 SeitenLipidos y Acidos Nucleicos (Trabajo) - Del Aguila Soto, OgerÔger Ernesto D. Aguila SotoNoch keine Bewertungen
- LIPIDOSDokument14 SeitenLIPIDOSItzel palacios100% (1)
- LIPIDOS BIOQUIMICA U3Dokument24 SeitenLIPIDOS BIOQUIMICA U3viri93Noch keine Bewertungen
- Lípidos: clasificación, estructura y función biológicaDokument21 SeitenLípidos: clasificación, estructura y función biológicaRox KatheNoch keine Bewertungen
- 2015 - Teoría N 03 Lípidos-BGDokument101 Seiten2015 - Teoría N 03 Lípidos-BGCATALINA NIETONoch keine Bewertungen
- LípidosDokument5 SeitenLípidoswpinedoNoch keine Bewertungen
- MPS 0103 Bases Bilogicas de La ConductaDokument225 SeitenMPS 0103 Bases Bilogicas de La ConductaChuy CastañedaNoch keine Bewertungen
- Cuestionario de BioquimicaDokument12 SeitenCuestionario de BioquimicaFernanda Lugo DiazNoch keine Bewertungen
- Lipidos y Membranas Celulares Parte IDokument43 SeitenLipidos y Membranas Celulares Parte IMaria AraujoNoch keine Bewertungen
- Lípidos: funciones, clasificación y estructura de fosfolípidosDokument18 SeitenLípidos: funciones, clasificación y estructura de fosfolípidosAdelis RuizNoch keine Bewertungen
- LípidosDokument3 SeitenLípidosnormanNoch keine Bewertungen
- Informe Nº6-Fabricación de JabónDokument24 SeitenInforme Nº6-Fabricación de JabónJesusNoch keine Bewertungen
- Lípidos esencialesDokument4 SeitenLípidos esencialestito1256Noch keine Bewertungen
- Informe de LipoproteínasDokument16 SeitenInforme de LipoproteínasFiorelaMHNoch keine Bewertungen
- LIPIDOS2020Dokument100 SeitenLIPIDOS2020Mohamed AléNoch keine Bewertungen
- LIPIDOSDokument4 SeitenLIPIDOSlmgarcia204Noch keine Bewertungen
- LipidosDokument17 SeitenLipidosGhia Valeriy Nahiara Sandoval ChilconNoch keine Bewertungen
- LipidosDokument10 SeitenLipidosElizabeth Quispe FloresNoch keine Bewertungen
- 3B PérezG LípidosDokument19 Seiten3B PérezG LípidosOscarZap359elproNoch keine Bewertungen
- Tema 4. Los LipidosDokument10 SeitenTema 4. Los LipidosSofiaAisaNoch keine Bewertungen
- Recien Nacido Sano UlisesDokument28 SeitenRecien Nacido Sano UlisesGisela PérezNoch keine Bewertungen
- Cuerpos Extraños en El EsófagoDokument8 SeitenCuerpos Extraños en El EsófagoGisela PérezNoch keine Bewertungen
- Síndrome de Stevens-JohnsonDokument18 SeitenSíndrome de Stevens-JohnsonGisela PérezNoch keine Bewertungen
- Columna VertebralDokument10 SeitenColumna VertebralGisela PérezNoch keine Bewertungen
- Infecciones Transmitidas Por VectoresDokument2 SeitenInfecciones Transmitidas Por VectoresGisela PérezNoch keine Bewertungen
- T 07 Hongos 142Dokument27 SeitenT 07 Hongos 142Gisela PérezNoch keine Bewertungen
- Compuestos CíclicosDokument9 SeitenCompuestos CíclicosGeometria Chimbotano Manuel LunaNoch keine Bewertungen
- Haluros de ÁcidoDokument6 SeitenHaluros de ÁcidoKarla DelgadoNoch keine Bewertungen
- Reacciones de adición al grupo carboniloDokument77 SeitenReacciones de adición al grupo carboniloMaría Alejandra GuerreroNoch keine Bewertungen
- Copia de Informe - Alcoholes y FenolesDokument14 SeitenCopia de Informe - Alcoholes y FenolesBreyner SanchezNoch keine Bewertungen
- ETERESDokument5 SeitenETERESVivianaPerdomo0% (1)
- Acido CarboxilicoDokument4 SeitenAcido CarboxilicoIdalfi Mármol PérezNoch keine Bewertungen
- Actividad Quimica 1Dokument9 SeitenActividad Quimica 1CARMEN ELINA MARTINEZNoch keine Bewertungen
- AOCS Ca 5a - 40 ACIDEZDokument5 SeitenAOCS Ca 5a - 40 ACIDEZHarold Isai Silvestre Gómez100% (1)
- Guia de Ejercicios Nomenclatura de HidrocarburosDokument10 SeitenGuia de Ejercicios Nomenclatura de HidrocarburosCarlosArnoldoManzanoAvelloNoch keine Bewertungen
- Sales neutras e hidrácidos: evaluación de fisicoquímicaDokument2 SeitenSales neutras e hidrácidos: evaluación de fisicoquímicaPelaitay Leonel100% (1)
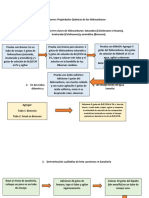
- Diagrama de Flujo Propiedades Químicas de Los HidrocarburosDokument2 SeitenDiagrama de Flujo Propiedades Químicas de Los HidrocarburosSantiago LopezNoch keine Bewertungen
- Grupos FuncionalesDokument9 SeitenGrupos Funcionalesfreddo800Noch keine Bewertungen
- Presentacion Formulacion 02-12-13Dokument87 SeitenPresentacion Formulacion 02-12-13Juana Rodriguez FloresNoch keine Bewertungen
- Nomenclatura de Compuestos Organicos PDFDokument34 SeitenNomenclatura de Compuestos Organicos PDFpepegrillo20Noch keine Bewertungen
- Tabla de IRDokument3 SeitenTabla de IRsociedad de servicios farmaceuticos a la comunidadNoch keine Bewertungen
- Acidos CarboxilicosDokument24 SeitenAcidos CarboxilicossiriusdarkNoch keine Bewertungen
- AmidasDokument9 SeitenAmidasJesus MurilloNoch keine Bewertungen
- La Importancia y de Utilidad de Los Distintos Tipos de Ácidos OxácidosDokument3 SeitenLa Importancia y de Utilidad de Los Distintos Tipos de Ácidos OxácidosMaria vzqNoch keine Bewertungen
- FQ1B Pizarras Formula FormulaciónOrgánicaDokument17 SeitenFQ1B Pizarras Formula FormulaciónOrgánicaMiguel El RuSsNoch keine Bewertungen
- BCM HidrocarburosDokument14 SeitenBCM HidrocarburosAndres ToscanoNoch keine Bewertungen
- Hidrocarburos SaturadosDokument7 SeitenHidrocarburos SaturadosRenato Proleon100% (1)
- Hidrocarburos Insaturados Alquenos y AlquinosDokument9 SeitenHidrocarburos Insaturados Alquenos y AlquinosAlexander Pernia DiazNoch keine Bewertungen
- Curvas PatronDokument55 SeitenCurvas PatronAna TorrealbaNoch keine Bewertungen
- Diapositivas Quimica OrganicaDokument37 SeitenDiapositivas Quimica OrganicaGilberthAlvaCelmiNoch keine Bewertungen
- Nomenclatura Química - Profesora Gabriela MartínezDokument8 SeitenNomenclatura Química - Profesora Gabriela MartínezmiaNoch keine Bewertungen
- Acidos Carboxilicos y DerivadosDokument12 SeitenAcidos Carboxilicos y DerivadosEliasDaniel Pacaya PanaifoNoch keine Bewertungen
- 2014guia de Problemas Nº5Dokument2 Seiten2014guia de Problemas Nº5jagus210% (2)
- Nomenclatura de oxoácidos: prefijos y sufijos para ácidos de no metales y metales de transiciónDokument2 SeitenNomenclatura de oxoácidos: prefijos y sufijos para ácidos de no metales y metales de transiciónFlavio CanterosNoch keine Bewertungen
- Haluro de AlquiloDokument4 SeitenHaluro de AlquiloAmneris Adriana VelàsquezNoch keine Bewertungen
- BecenosDokument6 SeitenBecenosAlex Gutierrez MarquezNoch keine Bewertungen