Das könnte Ihnen auch gefallen
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeVon EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeBewertung: 4 von 5 Sternen4/5 (5795)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreVon EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreBewertung: 4 von 5 Sternen4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItVon EverandNever Split the Difference: Negotiating As If Your Life Depended On ItBewertung: 4.5 von 5 Sternen4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceVon EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceBewertung: 4 von 5 Sternen4/5 (895)
- Grit: The Power of Passion and PerseveranceVon EverandGrit: The Power of Passion and PerseveranceBewertung: 4 von 5 Sternen4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeVon EverandShoe Dog: A Memoir by the Creator of NikeBewertung: 4.5 von 5 Sternen4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersVon EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersBewertung: 4.5 von 5 Sternen4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureVon EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureBewertung: 4.5 von 5 Sternen4.5/5 (474)
- Her Body and Other Parties: StoriesVon EverandHer Body and Other Parties: StoriesBewertung: 4 von 5 Sternen4/5 (821)
- The Emperor of All Maladies: A Biography of CancerVon EverandThe Emperor of All Maladies: A Biography of CancerBewertung: 4.5 von 5 Sternen4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Von EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Bewertung: 4.5 von 5 Sternen4.5/5 (121)
- The Little Book of Hygge: Danish Secrets to Happy LivingVon EverandThe Little Book of Hygge: Danish Secrets to Happy LivingBewertung: 3.5 von 5 Sternen3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyVon EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyBewertung: 3.5 von 5 Sternen3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)Von EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Bewertung: 4 von 5 Sternen4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaVon EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaBewertung: 4.5 von 5 Sternen4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryVon EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryBewertung: 3.5 von 5 Sternen3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnVon EverandTeam of Rivals: The Political Genius of Abraham LincolnBewertung: 4.5 von 5 Sternen4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealVon EverandOn Fire: The (Burning) Case for a Green New DealBewertung: 4 von 5 Sternen4/5 (74)
- The Unwinding: An Inner History of the New AmericaVon EverandThe Unwinding: An Inner History of the New AmericaBewertung: 4 von 5 Sternen4/5 (45)
- Lee Asher Card Magic Like The Asher Twist, PDFDokument1 SeiteLee Asher Card Magic Like The Asher Twist, PDFNebojša JoksimovićNoch keine Bewertungen
- BYO - Wine Maker - Beginner's Guide To Win - 2004 PDFDokument46 SeitenBYO - Wine Maker - Beginner's Guide To Win - 2004 PDFNebojša JoksimovićNoch keine Bewertungen
- Straw-Bale Building - Home Power, IncDokument6 SeitenStraw-Bale Building - Home Power, IncNebojša JoksimovićNoch keine Bewertungen
- Tele Standard PDFDokument1 SeiteTele Standard PDFNebojša JoksimovićNoch keine Bewertungen
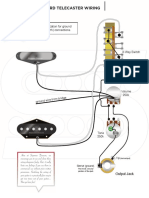
- Standard Telecaster Wiring: Location For Ground (Earth) ConnectionsDokument1 SeiteStandard Telecaster Wiring: Location For Ground (Earth) ConnectionsNebojša JoksimovićNoch keine Bewertungen
- Mustang With 2 Single Coil Sized Humbuckers: Red & White Wires Get Soldered Together and Taped OffDokument1 SeiteMustang With 2 Single Coil Sized Humbuckers: Red & White Wires Get Soldered Together and Taped OffNebojša JoksimovićNoch keine Bewertungen
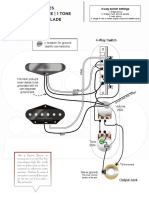
- 2 Singles 1 Volume - 1 Tone 4 Way Blade: Location For Ground (Earth) ConnectionsDokument1 Seite2 Singles 1 Volume - 1 Tone 4 Way Blade: Location For Ground (Earth) ConnectionsNebojša JoksimovićNoch keine Bewertungen
- 5 Minute Practice PDFDokument1 Seite5 Minute Practice PDFNebojša JoksimovićNoch keine Bewertungen
- 5 Minute Practice PDFDokument1 Seite5 Minute Practice PDFNebojša JoksimovićNoch keine Bewertungen
- 5 Minute Practice PDFDokument1 Seite5 Minute Practice PDFNebojša JoksimovićNoch keine Bewertungen
- 400GSV PDFDokument1 Seite400GSV PDFNebojša JoksimovićNoch keine Bewertungen
- Weight (With Mounting Hardware) : 0.2 Oz TEL:607-532-9404 FAX:607-532-9530Dokument1 SeiteWeight (With Mounting Hardware) : 0.2 Oz TEL:607-532-9404 FAX:607-532-9530Nebojša JoksimovićNoch keine Bewertungen
- 5 Minute Practice PDFDokument1 Seite5 Minute Practice PDFNebojša JoksimovićNoch keine Bewertungen
- Whipper Clipper: Bill of Materials Resistors CapacitorsDokument5 SeitenWhipper Clipper: Bill of Materials Resistors CapacitorsNebojša JoksimovićNoch keine Bewertungen
- Whipped Llama: Design by Erik VincentDokument6 SeitenWhipped Llama: Design by Erik VincentNebojša JoksimovićNoch keine Bewertungen
- 2 Singles 1 Volume - 1 Tone 4 Way Blade: Location For Ground (Earth) ConnectionsDokument1 Seite2 Singles 1 Volume - 1 Tone 4 Way Blade: Location For Ground (Earth) ConnectionsNebojša JoksimovićNoch keine Bewertungen
- AJINOMOTO 2013 Ideal Amino Acid Profile For PigletsDokument28 SeitenAJINOMOTO 2013 Ideal Amino Acid Profile For PigletsFreddy Alexander Horna Morillo100% (1)
- Leadership Roles and Management Functions in Nursing Theory and ApplicationDokument2 SeitenLeadership Roles and Management Functions in Nursing Theory and Applicationivan0% (3)
- Marpappa EASADokument5 SeitenMarpappa EASAAshley SamNoch keine Bewertungen
- Brosura Oxyhelp PDFDokument12 SeitenBrosura Oxyhelp PDFAftab Naseem100% (1)
- Paramagnetic Article PDFDokument5 SeitenParamagnetic Article PDFJonathan SinclairNoch keine Bewertungen
- Cameron Scanner 2000 Hardware User Manual PDFDokument11 SeitenCameron Scanner 2000 Hardware User Manual PDFPes SilvaNoch keine Bewertungen
- BRSM Form 009 - QMS MDD TPDDokument15 SeitenBRSM Form 009 - QMS MDD TPDAnonymous q8lh3fldWMNoch keine Bewertungen
- Manuscript 1Dokument26 SeitenManuscript 1Juan Paolo CapatiNoch keine Bewertungen
- Achai, Sydney Jill S. GE 15 - SIM - ULOcDokument13 SeitenAchai, Sydney Jill S. GE 15 - SIM - ULOcSydney AchaiNoch keine Bewertungen
- Competency Competency Multiple Choice Multiple Choice ComputationDokument4 SeitenCompetency Competency Multiple Choice Multiple Choice ComputationAaron James LicoNoch keine Bewertungen
- Cable Selection Table For CapacitorDokument1 SeiteCable Selection Table For CapacitorShashiSharmaNoch keine Bewertungen
- DENSO Diagnostic TipsDokument1 SeiteDENSO Diagnostic TipsVerona MamaiaNoch keine Bewertungen
- Permatex, Inc - Ultra Gasket Sealant 1ozDokument3 SeitenPermatex, Inc - Ultra Gasket Sealant 1ozjaredf@jfelectric.comNoch keine Bewertungen
- 2016 Liberal Policy ResolutionsDokument86 Seiten2016 Liberal Policy ResolutionsCPAC TVNoch keine Bewertungen
- Basic PreservationDokument14 SeitenBasic Preservationrovinj1Noch keine Bewertungen
- Thai Cuisine: Reporters: Bantayan, Kenneth Samejon, Clarish Lovely Relevo, Mary GraceDokument47 SeitenThai Cuisine: Reporters: Bantayan, Kenneth Samejon, Clarish Lovely Relevo, Mary Gracemiralona relevoNoch keine Bewertungen
- Bradycardia AlgorithmDokument1 SeiteBradycardia AlgorithmGideon BahuleNoch keine Bewertungen
- Fire CodeDokument1 SeiteFire CodeShainette VillarazaNoch keine Bewertungen
- Amirtha ProjectDokument18 SeitenAmirtha Projectaeriel judson100% (1)
- TEC3000 Series On/Off or Floating Fan Coil and Individual Zone Thermostat Controllers With Dehumidification CapabilityDokument48 SeitenTEC3000 Series On/Off or Floating Fan Coil and Individual Zone Thermostat Controllers With Dehumidification CapabilityIvar Denicia DelgadilloNoch keine Bewertungen
- Bituminous MixesDokument13 SeitenBituminous MixesRanjit SinghNoch keine Bewertungen
- Granulation Process Basic UnderstandingDokument3 SeitenGranulation Process Basic UnderstandingRainMan75Noch keine Bewertungen
- Guide For Visual Inspection of Structural Concrete Building ComponentsDokument76 SeitenGuide For Visual Inspection of Structural Concrete Building ComponentsMazin AlwashNoch keine Bewertungen
- digiPHONENT UG enDokument44 SeitendigiPHONENT UG enIrving Javier Leal OrtizNoch keine Bewertungen
- Arcelor Mittal Operations: Operational Area Is Sub-Divided Into 4 PartsDokument5 SeitenArcelor Mittal Operations: Operational Area Is Sub-Divided Into 4 Partsarpit agrawalNoch keine Bewertungen
- Peritoneal Dialysis Unit Renal Department SGH PD WPI 097 Workplace InstructionDokument10 SeitenPeritoneal Dialysis Unit Renal Department SGH PD WPI 097 Workplace InstructionAjeng SuparwiNoch keine Bewertungen
- Flaxseed Paper PublishedDokument4 SeitenFlaxseed Paper PublishedValentina GarzonNoch keine Bewertungen
- Medical Devices Industry in IndiaDokument6 SeitenMedical Devices Industry in IndiaMurali Krishna Reddy100% (1)
- Tokyo Fact SheetDokument17 SeitenTokyo Fact Sheethoangnguyen2401Noch keine Bewertungen
- Vertico SynchroDokument16 SeitenVertico SynchrozpramasterNoch keine Bewertungen